The gene's-eye view of quantitative genetics
Pith reviewed 2026-05-23 02:54 UTC · model grok-4.3
The pith
As the number of loci tends to infinity under strong recombination, each locus evolves according to a McKean-Vlasov SDE whose selection term depends on the law of the focal locus itself.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
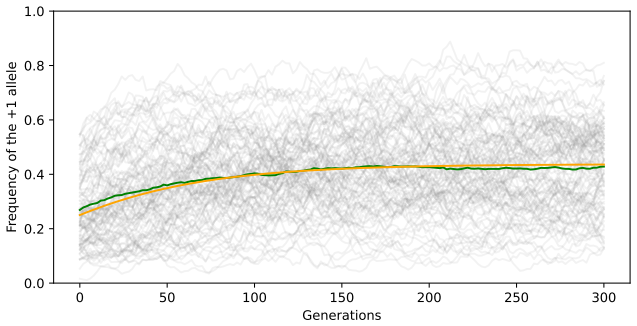
In the infinite-loci limit under strong recombination, the evolution of the allelic frequency at a typical locus is described by a McKean-Vlasov SDE whose drift term depends on the law of the process itself, together with the associated Fokker-Planck integro-partial differential equation. This mean-field interaction implies that two distinct loci evolve independently, and under suitable fitness functions the stationary distribution of allele frequencies at a locus can be written explicitly.
What carries the argument
The McKean-Vlasov stochastic differential equation for the allelic frequency process at a focal locus, whose coefficients are functionals of the marginal law of that process, arising from the mean-field limit of the finite-loci model.
If this is right
- Distinct loci evolve independently in the limit.
- Explicit stationary distributions for allelic frequencies exist under assumptions on the fitness function.
- The overall trait distribution evolves according to the common marginal law of any single locus.
- Selection at each locus is fully determined by the population-level average allelic behavior rather than by specific pairwise interactions.
Where Pith is reading between the lines
- The same mean-field limit could supply effective single-locus approximations for large-scale genomic data without requiring an assumed shape for the trait distribution.
- The independence result implies that linkage disequilibrium between loci decays to zero under strong recombination even in the presence of selection.
- Numerical simulation of the original finite-loci model for moderately large numbers of loci could test how quickly the mean-field description becomes accurate.
Load-bearing premise
Strong recombination must hold when the number of loci tends to infinity, so that the effect of all other loci on a focal locus can be replaced by the average law of the focal locus.
What would settle it
Persistent statistical dependence between allelic frequencies at two distinct loci in a large-loci system with strong recombination would contradict the claimed independence.
Figures
read the original abstract
Modelling the evolution of a continuous trait in a biological population is one of the oldest problems in evolutionary biology, which led to the birth of quantitative genetics. With the recent development of GWAS methods, it has become essential to link the evolution of the trait distribution to the underlying evolution of allelic frequencies at many loci, co-contributing to the trait value. The way most articles go about this is to make assumptions on the trait distribution, and use Wright's formula to model how the evolution of the trait translates on each individual locus. Here, we take a gene's eye-view of the system, starting from an explicit finite-loci model with selection, drift, recombination and mutation, in which the trait value is a direct product of the genome. We let the number of loci go to infinity under the assumption of strong recombination, and characterize the limit behavior of a given locus with a McKean-Vlasov SDE and the corresponding Fokker-Planck IPDE. In words, the selection on a typical locus depends on the mean behaviour of the other loci which can be approximated with the law of the focal locus. Results include the independence of two loci and explicit stationary distribution for allelic frequencies at a given locus (under some assumptions on the fitness function).
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript starts from an explicit finite-loci model incorporating selection, drift, recombination and mutation, with the trait value determined directly by the genome. It passes to the limit of infinitely many loci under a strong-recombination assumption and characterizes the limiting behavior of a typical locus by a McKean-Vlasov SDE whose drift depends on the law of the process itself; the associated nonlinear Fokker-Planck IPDE is derived. The results include asymptotic independence of distinct loci and, under further assumptions on the fitness function, an explicit stationary distribution for allelic frequencies at a focal locus.
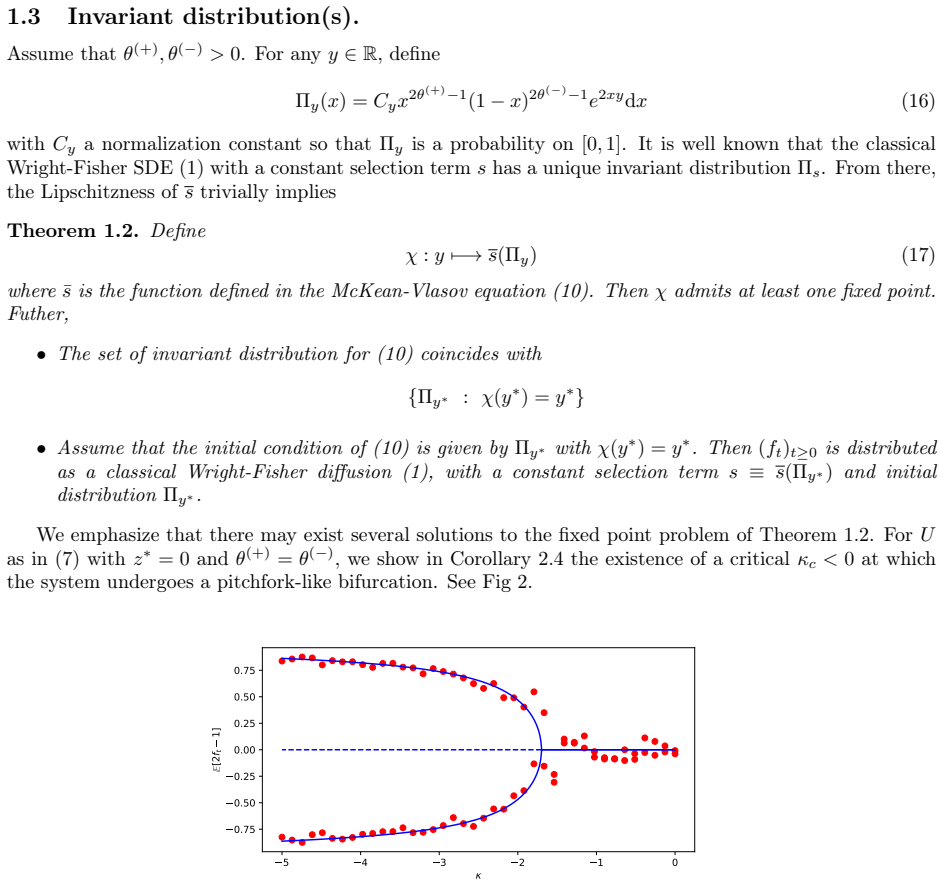
Significance. If the limit construction is made rigorous, the work supplies a mathematically grounded gene's-eye perspective that links finite-locus dynamics to mean-field approximations in quantitative genetics. The McKean-Vlasov formulation and the explicit stationary distributions (when available) constitute concrete, falsifiable outputs that could be compared with GWAS data or simulation.
major comments (1)
- [limit procedure / abstract] The limit procedure (abstract and the construction leading to the McKean-Vlasov SDE) invokes “strong recombination” to obtain mean-field closure, yet supplies no scaling of the per-locus recombination rate with the number of loci N. Without a rate that diverges sufficiently rapidly with N, residual linkage disequilibrium of order 1/N or slower can persist, so that the empirical measure of the remaining loci need not converge to the law of the focal locus; this directly affects the validity of the nonlinear drift, the independence statement, and the Fokker-Planck IPDE.
minor comments (1)
- The abstract states that an explicit stationary distribution is obtained “under some assumptions on the fitness function,” but does not list those assumptions; stating them explicitly near the statement of the stationary result would improve clarity.
Simulated Author's Rebuttal
We thank the referee for their careful reading of the manuscript and for highlighting an important point regarding the limit procedure. We address the comment below.
read point-by-point responses
-
Referee: [limit procedure / abstract] The limit procedure (abstract and the construction leading to the McKean-Vlasov SDE) invokes “strong recombination” to obtain mean-field closure, yet supplies no scaling of the per-locus recombination rate with the number of loci N. Without a rate that diverges sufficiently rapidly with N, residual linkage disequilibrium of order 1/N or slower can persist, so that the empirical measure of the remaining loci need not converge to the law of the focal locus; this directly affects the validity of the nonlinear drift, the independence statement, and the Fokker-Planck IPDE.
Authors: The referee correctly identifies that the manuscript does not provide an explicit scaling for the recombination rate in terms of N. In the current version, 'strong recombination' is used qualitatively to indicate that recombination is fast enough to justify the mean-field approximation. To rigorously justify the convergence to the McKean-Vlasov SDE and the asymptotic independence, a specific scaling condition is required. We agree that this needs to be addressed and will revise the manuscript to include a precise assumption, for example requiring the recombination rate r_N to satisfy r_N / log N -> infinity or an appropriate condition that ensures LD decays sufficiently fast. This revision will be made in the abstract, introduction, and the statement of the main results, along with a discussion of why this scaling suffices for the limit. revision: yes
Circularity Check
No circularity: explicit finite-loci model to mean-field limit
full rationale
The derivation begins with an explicit finite-loci model incorporating selection, drift, recombination and mutation, then passes to the N→∞ limit under a strong-recombination assumption to obtain a McKean-Vlasov SDE whose drift depends on the law of the focal locus. No step reduces by construction to its own inputs, no parameters are fitted on a subset and re-labeled as predictions, and no load-bearing self-citations or imported uniqueness theorems appear. The procedure is a standard probabilistic limit argument whose validity is external to the target stationary distributions or independence statements.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption Strong recombination assumption in the infinite loci limit
- domain assumption Existence of the limit as number of loci goes to infinity
Reference graph
Works this paper leans on
-
[1]
Polygenic adaptation: From sweeps to subtle frequency shifts.PLoS genetics, 15(3):e1008035, 2019
Ilse Höllinger, Pleuni S Pennings, and Joachim Hermisson. Polygenic adaptation: From sweeps to subtle frequency shifts.PLoS genetics, 15(3):e1008035, 2019
work page 2019
-
[2]
Y B. Simons, K Bullaughey, R R. Hudson, and G Sella. A population genetic interpretation of gwas findings for human quantitative traits.PLoS Biology, 16:e2002985, 4 2017
work page 2017
-
[3]
R A Fisher. The correlation between relatives on the supposition of mendelian inheritance.Transaction Royal Society of Edinburgh, 52:399–433, 1918
work page 1918
-
[4]
Francis Galton. Regression towards mediocrity in hereditary stature.The Journal of the Anthropological Institute of Great Britain and Ireland, 15:246–263, 1886
-
[5]
The infinitesimal model: Definition, derivation, and implications
N H Barton, A M Etheridge, and A Véber. The infinitesimal model: Definition, derivation, and implications. Theoretical Population Biology, 118:50–73, 12 2017
work page 2017
-
[6]
B Walsh and M Lynch.Evolution and selection of quantitative traits. Oxford University Press, 2018
work page 2018
-
[7]
Natural selection and random genetic drift in phenotypic evolution
Russell Lande. Natural selection and random genetic drift in phenotypic evolution. Evolution, 30(2):314–334, 06 1976
work page 1976
-
[8]
Léonard Dekens. Quantifying the influence of sexual reproduction on the spatial eco-evo dynamics of species characterized by complex traits: asymptotic methods. PhD thesis, Université de Lyon, 2022
work page 2022
-
[9]
A computational view of population genetics
Yuval Rabani, Yuri Rabinovich, and Alistair Sinclair. A computational view of population genetics. In Proceedings of the twenty-seventh annual ACM symposium on Theory of computing, pages 83–92, 1995
work page 1995
-
[10]
Servet Martínez. A probabilistic analysis of a discrete-time evolution in recombination.Advances in Applied Mathematics, 91:115–136, 2017. 36
work page 2017
-
[11]
Ellen Baake, Michael Baake, and Majid Salamat. The general recombination equation in continuous time and its solution.Discrete and Continuous Dynamical Systems, 36(1):63–95, 2016
work page 2016
-
[12]
An exactly solved model for mutation, recombination and selection
Michael Baake and Ellen Baake. An exactly solved model for mutation, recombination and selection. Canadian Journal of Mathematics, 55(1):3–41, 2003
work page 2003
-
[13]
Entropy production in nonlinear recombination models
P Caputo and A Sinclair. Entropy production in nonlinear recombination models. Bernoulli, 24(4B):3246 – 3282, 2018
work page 2018
-
[14]
American Mathematical Society, 2013
Steven N Evans, David Steinsaltz, and Kenneth Wachter.A mutation-selection model with recombina- tion for general genotypes, volume 222. American Mathematical Society, 2013
work page 2013
-
[15]
George R. Price. Selection and covariance.Nature, 227(5257):520–521, Aug 1970
work page 1970
-
[16]
Sabin Lessard. Fisher’s fundamental theorem of natural selection revisited.Theoretical Population Biology, 52(2):119–136, 1997
work page 1997
-
[17]
Matthias Steinrücken, YX Rachel Wang, and Yun S Song. An explicit transition density expansion for a multi-allelic wright–fisher diffusion with general diploid selection.Theoretical population biology, 83:1–14, 2013
work page 2013
-
[18]
Aclassofdegeneratediffusionprocessesoccurringinpopulationgenetics
S.N.Ethier. Aclassofdegeneratediffusionprocessesoccurringinpopulationgenetics. Communications on Pure and Applied Mathematics, 29(5):483–493, 1976
work page 1976
-
[19]
Olivier Tenaillon. The utility of fisher’s geometric model in evolutionary genetics.Annual review of ecology, evolution, and systematics, 45:179–201, 2014
work page 2014
-
[20]
Propagation of chaos: A review of models, methods and applications
LP Chaintron and A Diez. Propagation of chaos: A review of models, methods and applications. i. models and methods.Kinetic and Related Models, 15(6):895–1015, 2022
work page 2022
-
[21]
Patrick Billingsley.Convergence of probability measures. John Wiley & Sons, 1968
work page 1968
-
[22]
M. G. Bulmer. Linkage disequilibrium and genetic variability.Genetical Research, 23(3):281–289, 1974
work page 1974
-
[23]
Polygenicadaptationafterasuddenchangeinenvironment
LKHaywardandGSella. Polygenicadaptationafterasuddenchangeinenvironment. eLife, 11:e66697, sep 2022
work page 2022
-
[24]
Guy Sella and Nicholas H. Barton. Thinking about the evolution of complex traits in the era of genome-wide association studies. Annual Review of Genomics and Human Genetics, 20:461–493, 8 2019
work page 2019
-
[25]
BDH Latter. Natural selection for an intermediate optimum.Australian Journal of Biological Sciences, 13(1):30–35, 1960
work page 1960
-
[26]
M. G. Bulmer. The genetic variability of polygenic characters under optimizing selection, mutation and drift. Genetical Research, 19(1):17–25, 1972
work page 1972
-
[27]
J. Avid Ågren. Sewall wright’s criticism of the gene’s-eye view of evolution.Evolution, 75, 10 2021
work page 2021
-
[28]
Dynamic maximum entropy provides accurate approximation of structured population dynamics
Katarína Bod’ová, Enikő Szép, and Nicholas H Barton. Dynamic maximum entropy provides accurate approximation of structured population dynamics. PLoS Computational Biology, 17(12):e1009661, 2021
work page 2021
-
[29]
Armita Nourmohammad, Stephan Schiffels, and Michael Lässig. Evolution of molecular phenotypes under stabilizing selection.Journal of Statistical Mechanics: Theory and Experiment, 2013(01):P01012, jan 2013
work page 2013
-
[30]
Todd L. Parsons and Peter L. Ralph. Large effects and the infinitesimal model.Theoretical Population Biology, 156:117–129, 2024
work page 2024
-
[31]
The effect of selection against extreme deviants based on deviation or on homozygosis
Alan Robertson. The effect of selection against extreme deviants based on deviation or on homozygosis. Journal of Genetics, 54:236–248, 1956
work page 1956
-
[32]
Michael Lynch. The lower bound to the evolution of mutation rates.Genome Biology and Evolution, 3:1107–1118, 08 2011
work page 2011
-
[33]
Kevin J. Dawson. Evolutionarily stable mutation rates.Journal of Theoretical Biology, 194(1):143–157, 1998. 37
work page 1998
-
[34]
V Dichio, H-L Zeng, and E Aurell. Statistical genetics in and out of quasi-linkage equilibrium.Reports on Progress in Physics, 86(5):052601, apr 2023
work page 2023
-
[35]
Springer Science & Business Media, 1997
D W Stroock and S R S Varadhan.Multidimensional diffusion processes, volume 233. Springer Science & Business Media, 1997
work page 1997
-
[36]
Well-posedness of multidimensional diffusion processes with weakly differentiable co- efficients
Dario Trevisan. Well-posedness of multidimensional diffusion processes with weakly differentiable co- efficients. Electronic Journal of Probability, 21(none):1 – 41, 2016
work page 2016
-
[37]
Yu I Lyubich. Basic concepts and theorems of the evolutionary genetics of free populations.Russian Mathematical Surveys, 26(5):51, oct 1971
work page 1971
-
[38]
Solutions of a stochastic differential equation forced onto a manifold by a large drift
Gary Shon Katzenberger. Solutions of a stochastic differential equation forced onto a manifold by a large drift. The University of Wisconsin-Madison, 1990
work page 1990
-
[39]
SN Ethier. A limit theorem for two-locus diffusion models in population genetics.Journal of Applied Probability, 16(2):402–408, 1979
work page 1979
-
[40]
WENDELL H. FLEMING and MICHEL VIOT. Some measure-valued markov processes in population genetics theory.Indiana University Mathematics Journal, 28(5):817–843, 1979
work page 1979
-
[41]
Ellen Baake and Michael Baake. Haldane linearisation done right: Solving the nonlinear recombination equation the easy way.Discrete and Continuous Dynamical Systems, 36(12):6645–6656, 2016
work page 2016
-
[42]
Propagation of chaos: A review of models, methods and applications
LP Chaintron and A Diez. Propagation of chaos: A review of models, methods and applications. ii. applications. Kinetic and Related Models, 15(6):1017–1173, 2022
work page 2022
-
[43]
Sylvie Roelly-Coppoletta. A criterion of convergence of measure-valued processes: application to mea- sure branching processes.Stochastics: An International Journal of Probability and Stochastic Processes, 17(1-2):43–65, 1986
work page 1986
-
[44]
Continuous martingales and Brownian motion, volume 293
Daniel Revuz and Marc Yor. Continuous martingales and Brownian motion, volume 293. Springer Science & Business Media, 2013
work page 2013
- [45]
-
[46]
Henry J Landau and Lawrence A Shepp. On the supremum of a gaussian process.Sankhy¯ a: The Indian Journal of Statistics, Series A, pages 369–378, 1970. A Appendix A.1 The L1 law of the iterated logarithm Theorem A.1. There is a universal constantC > 0 such that for any a continuous martingale(Mt)t∈[0,T ) with 0 ≤ T ≤ +∞ we have E sup t∈[0,T ) Mtr 1 +...
work page 1970
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.