Viability Space Decomposition: A geometric partition of survival outcomes in single- and multi-agent systems
Pith reviewed 2026-05-19 19:50 UTC · model grok-4.3
The pith
Viability space decomposition partitions state space into regions of qualitatively similar survival outcomes using mortality, ordering, and collapse manifolds.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Viability space decomposition supplies a geometric partition of the state space for viability-constrained ODE models by identifying mortality manifolds that separate viable from non-viable states, ordering manifolds that rank successive viability limits, and collapse manifolds that resolve multi-agent interactions, thereby yielding a complete viability portrait of qualitatively distinct survival outcomes.
What carries the argument
Viability space decomposition, which uses mortality, ordering, and collapse manifolds to produce a viability portrait that partitions state space according to survival outcomes.
Load-bearing premise
The introduced manifolds can be identified and combined to achieve a complete decomposition of all survival outcomes for the full class of viability-constrained ODE models without leaving regions unclassified.
What would settle it
A viability-constrained ODE model in which some open regions of state space exhibit mixed or unclassified survival outcomes after all mortality, ordering, and collapse manifolds have been located.
Figures









read the original abstract
What determines whether an organism or collective will survive under particular conditions? This question is asked across the life sciences when determining adaptive fit, developing efficacious treatments for diseases, and assessing the risks posed by ecological shifts. To aid their investigations, researchers employ models of agents which must respect particular constraints to remain alive. By constraining the dynamics of these agents to bounded viability regions, these models form a class of extended dynamical systems where transient dynamics can lead to death, making traditional attractors and separatrices insufficient for characterizing the global space of possible behaviors. To remedy this, we develop viability space decomposition, an analysis framework for ordinary differential equation models of agents with viability constraints. We first introduce the general theory, revealing how several new classes of manifolds (mortality, ordering, and collapse) permit a complete decomposition of state space into regions of qualitatively similar survival outcomes: a viability portrait. We then demonstrate the method by completely analyzing the global behavior of three models: a subcellular network, a behaving cell with the same physiology, and two coupled cell networks. Finally, we finish by discussing how the framework scales and future directions for its development and application.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper develops viability space decomposition, an analysis framework for ODE models of agents subject to viability constraints. It introduces mortality, ordering, and collapse manifolds that are claimed to enable a complete geometric partition of state space into regions of qualitatively similar survival outcomes (a viability portrait). The framework is demonstrated via full global analysis of three models: a subcellular network, a single behaving cell, and two coupled cells.
Significance. If the completeness claim holds, the work supplies a systematic geometric tool for characterizing global behavior in viability-constrained dynamical systems, where traditional attractors and separatrices are insufficient. The three explicit demonstrations provide concrete, reproducible examples of the method in action and could support falsifiable predictions about survival regions in biological models.
major comments (1)
- [General theory] General theory (prior to the demonstrations): the manuscript asserts that the new manifolds permit a complete decomposition for the full class of viability-constrained ODE models, yet no explicit theorem or proof is supplied that guarantees the manifolds can always be identified and that their union (together with viability regions) exhausts state space without unclassified trajectories or overlaps. The demonstrations are restricted to three low-dimensional cases with relatively simple viability boundaries; this leaves the generality claim load-bearing but unproven for higher-dimensional or topologically complex systems.
minor comments (1)
- [Abstract] The abstract refers to 'several new classes of manifolds' without giving their defining properties or construction; a brief definitional sentence would improve immediate readability.
Simulated Author's Rebuttal
We thank the referee for their positive evaluation of the work's significance and for the constructive major comment. We address the concern regarding the general theory below.
read point-by-point responses
-
Referee: [General theory] General theory (prior to the demonstrations): the manuscript asserts that the new manifolds permit a complete decomposition for the full class of viability-constrained ODE models, yet no explicit theorem or proof is supplied that guarantees the manifolds can always be identified and that their union (together with viability regions) exhausts state space without unclassified trajectories or overlaps. The demonstrations are restricted to three low-dimensional cases with relatively simple viability boundaries; this leaves the generality claim load-bearing but unproven for higher-dimensional or topologically complex systems.
Authors: We agree that a formal theorem would strengthen the presentation of the general theory. In the revised manuscript we will add an explicit theorem stating that, under standard assumptions (Lipschitz continuity of the vector field and compactness of the viability region), the mortality, ordering, and collapse manifolds together with the viability regions form a partition of the state space with no unclassified trajectories or overlaps. The proof will proceed by showing that every initial condition has a unique forward orbit that either remains in the viability region for all time or intersects the viability boundary at a first time, after which the orbit is classified by the type of boundary intersection (mortality, ordering, or collapse). We will also include a brief discussion of the coordinate-free, dimension-independent nature of the construction, noting that while the three demonstrations are low-dimensional for computational clarity, the definitions and partition property extend directly to higher-dimensional and topologically more complex viability sets. We will add a remark on practical identification challenges in high dimensions without altering the theoretical completeness claim. revision: yes
Circularity Check
No significant circularity; framework introduces independent geometric constructs
full rationale
The paper defines viability space decomposition by introducing new manifold classes (mortality, ordering, collapse) as part of a general theory for viability-constrained ODEs, then applies them to three specific low-dimensional models. No step reduces a claimed prediction or completeness result to a fitted parameter, self-citation chain, or definitional renaming; the central decomposition is constructed explicitly from the stated geometric partitions rather than presupposing the target classification. Demonstrations remain illustrative rather than load-bearing for the general claim, and the derivation stays self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Models of agents with viability constraints form extended dynamical systems where transient dynamics can lead to death and traditional attractors/separatrices are insufficient.
invented entities (1)
-
mortality manifolds, ordering manifolds, collapse manifolds
no independent evidence
Lean theorems connected to this paper
-
IndisputableMonolith/Foundation/AbsoluteFloorClosure.leanreality_from_one_distinction unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
mortality manifolds M = {ϕt(m) : t ≤ 0} ... ordering manifolds O ... collapse manifolds C
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
viability portrait ... hybrid automaton with guard conditions Gα
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
This makes the set of molecules autocatalytic
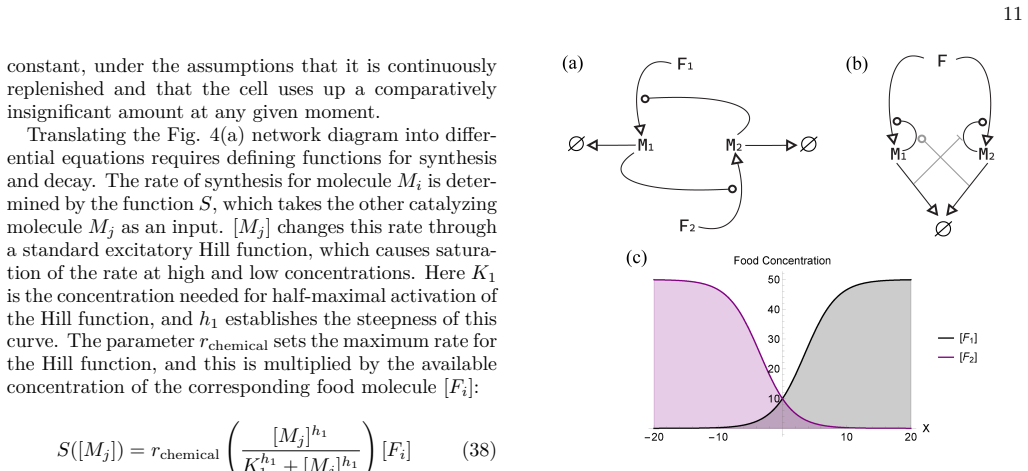
Model specification The single-cell physiology is composed of two molecu- lar species M = {M1, M2} which reciprocally catalyze each other and are synthesized from one of two food molecules F = {F1, F2} respectively. This makes the set of molecules autocatalytic. Fig. 4(a) depicts this cir- cuit with normal arrows representing material transfor- mation, ro...
-
[2]
Numerical exploration To approximate the possible existential outcomes in the single-cell physiology, we uniformly sample a million initial conditions in the range [Mi] ∈ [0, 20]. We then numerically integrate each initial condition for up to 800 arbitrary time units (ATU), and color the initial condi- tion according to whether it survives or violates one...
-
[3]
This reveals five distinct regions. The complement of the viability region V α, colored black, corresponds to where the total survival time is zero because the agent cannot exist beyond its viability region. The viability boundary ∂Vα trivially separates this portion from the other subsets where the agent persists for at least some time. Within the viabil...
-
[4]
Viability portrait A phase portrait analysis within the cell’s viability region reveals two equilibrium points: a saddle node EPS = (4 .008, 2.407) (light green point) and a sta- ble equilibrium point EPΩ = (13 .811, 4.686) (dark blue point), shown in Fig. 5(b). Sampling the flow, we can see that the asymptotically viable trajectories (green) con- verge t...
-
[5]
gives an ordering manifold O shown as a purple con- tour that separates T ′ 1 and T ′ 2 (Fig. 5(c)). Zooming in closer around o and sampling another 160,000 initial con- ditions for improved resolution, we can clearly see that O separates the two regions while the W u branch belongs to T ′ 2 (Fig. 5(d)). This final manifold finishes carving out the differ...
-
[6]
Model specification Our behaving cell model has the same physiology and viability constraints as the previous model, with the main difference being that we now allow the cell to move in a one-dimensional environment, X, which has opposing gradients of food concentrations, established by the fol- lowing equations and illustrated in Fig. 4(c): [F1] = 10 0.2...
-
[7]
One representative slice is shown in Fig
Numerical exploration As with the cell physiology in a fixed environment, we approximate the possible existential outcomes for the behaving cell by sampling a million initial con- ditions spanning [Mi] ∈ [0, 20] simulated for 200 ATU, except now for slices of the environment X = [−20, −15, −10, −5, 0, 5, 10, 15, 20]. One representative slice is shown in F...
-
[8]
Viability Portrait Similar to the protocell analysis without behavior, a phase portrait analysis within Vα reveals a stable equi- librium point EPΩ = (6 .434, 6.434, 0) (dark blue) and a saddle node EPS = (4.020, 4.020, 0) (light green) (Fig. 6(d)). The branches of the saddle node’s one-dimensional unstable manifold W u (red trajectories) either converge ...
-
[9]
Model specification Like the single-cell physiology model, our multi-agent model will be comprised of two cells, α = {α1, α2}, where [M1] and [M2] belong to each of these agents respectively (Fig. 4(b)). We assume that these molecules are synthe- sized from the same precursor food molecule, but beyond this the synthesis is the same as in Eqn. 38: S([Mi]) ...
-
[10]
Numerical exploration We sample a million initial conditions within the do- main of interaction of the two cells X and then simu- late their trajectories for 800 ATU, revealing six discon- nected regions of qualitatively distinct survival outcomes (Fig. 8(a)). Regions are given labels according to their sequence of asymptotically and transiently viable ou...
-
[11]
Viability portrait Decomposing our two-cell state space demands that we understand how our agents’ dynamics unfold within the intersection of their viability regions X . A phase por- trait analysis of the unified state space reveals a stable equilibrium point EPX Ω = (7.167, 4.707) (dark blue point) and a saddle node EPX S = (2 .571, 8.716) (light green) ...
-
[12]
E. Schrödinger, What Is Life? The Physical Aspect of the Living Cell ; with Mind and Matter & Autobiographical Sketches, Canto Classics (Cambridge University Press, Cambridge ; New York, 1992)
work page 1992
-
[13]
Phillips, Schrödinger’s What Is Life? at 75, Cell Sys- tems 12, 465 (2021)
R. Phillips, Schrödinger’s What Is Life? at 75, Cell Sys- tems 12, 465 (2021)
work page 2021
-
[14]
R. D. Beer and E. A. Di Paolo, The theoretical foun- dations of enaction: Precariousness, Biosystems 223, 104823 (2023)
work page 2023
-
[15]
M. Loreau, From Populations to Ecosystems: The- oretical Foundations for a New Ecological Synthesis , Monographs in Population Biology (Princeton University Press, Princeton, 2010)
work page 2010
-
[16]
C. McShaffrey, E. Agmon, and R. D. Beer, Matters of life and death in computational cell biology, npj Syst Biol Appl 10.1038/s41540-026-00718-y (2026)
-
[17]
R. I. Joh and J. S. Weitz, To Lyse or Not to Lyse: Transient-Mediated Stochastic Fate Determination in Cells Infected by Bacteriophages, PLoS Comput Biol 7, e1002006 (2011). 23
work page 2011
-
[18]
A. Ghaffarizadeh, R. Heiland, S. H. Friedman, S. M. Mu- menthaler, and P. Macklin, PhysiCell: An open source physics-based cell simulator for 3-D multicellular sys- tems, PLoS Comput Biol 14, e1005991 (2018)
work page 2018
-
[19]
C. McShaffrey and R. Beer, Maintaining Viability with Multiple Needs, in 2022 IEEE Symposium Series on Computational Intelligence (SSCI) (IEEE, Singapore, Singapore, 2022) pp. 1523–1528
work page 2022
-
[20]
W. R. Ashby, The Physical Origin of Adaptation by Trial and Error, The Journal of General Psychology 32, 13 (1945)
work page 1945
-
[21]
W. R. Ashby, Design for a Brain , 2nd ed. (Springer Netherlands, Dordrecht, 1960)
work page 1960
-
[22]
R. D. Beer, A dynamical systems perspective on agent- environment interaction, Artificial Intelligence 72, 173 (1995)
work page 1995
-
[23]
N. Virgo, M. Biehl, M. Baltieri, and M. Capucci, A “Good Regulator Theorem” for Embodied Agents, in AL- IFE 2025: Ciphers of Life: Proceedings of the Artificial Life Conference 2025 (Kyoto, Japan, 2025) p. 46
work page 2025
-
[24]
E. O. Voit, A systems-theoretical framework for health and disease: Inflammation and preconditioning from an abstract modeling point of view, Mathematical Bio- sciences 217, 11 (2009)
work page 2009
-
[25]
J. D. Davis, C. M. Kumbale, Q. Zhang, and E. O. Voit, Dynamical systems approaches to personalized medicine, Current Opinion in Biotechnology 58, 168 (2019)
work page 2019
-
[26]
X. E. Barandiaran and M. D. Egbert, Norm-Establishing and Norm-Following in Autonomous Agency, Artificial Life 20, 5 (2014)
work page 2014
- [27]
-
[28]
Y. Himeoka, S. A. Horiguchi, and T. J. Kobayashi, The- oretical basis for cell deaths, Phys. Rev. Research 6, 043217 (2024)
work page 2024
-
[29]
M. D. Egbert and J. Pérez-Mercader, Methods for Mea- suring Viability and Evaluating Viability Indicators, Ar- tificial Life 24, 106 (2018)
work page 2018
-
[30]
A. Kolchinsky and D. H. Wolpert, Semantic informa- tion, autonomous agency and non-equilibrium statistical physics, Interface Focus. 8, 20180041 (2018)
work page 2018
-
[31]
S. Bartlett, A. W. Eckford, M. Egbert, M. Lingam, A. Kolchinsky, A. Frank, and G. Ghoshal, Physics of Life: Exploring Information as a Distinctive Feature of Living Systems, PRX Life 3, 037003 (2025)
work page 2025
-
[32]
M. Baltieri and K. Suzuki, Mathematical approaches to the study of agents (2025)
work page 2025
-
[33]
H. J. Chiel and R. D. Beer, The brain has a body: Adap- tive behavior emerges from interactions of nervous sys- tem, body and environment, Trends in Neurosciences 20, 553 (1997)
work page 1997
-
[34]
K. Aihara and H. Suzuki, Theory of hybrid dynamical systems and its applications to biological and medical systems, Phil. Trans. R. Soc. A. 368, 4893 (2010)
work page 2010
-
[35]
M. G. Cortes, Y. Lin, L. Zeng, and G. Balázsi, From Bench to Keyboard and Back Again: A Brief History of Lambda Phage Modeling, Annu. Rev. Biophys. 50, 117 (2021)
work page 2021
-
[36]
J. Yuan and D. Ofengeim, A guide to cell death pathways, Nat Rev Mol Cell Biol 25, 379 (2024)
work page 2024
-
[37]
R. Abraham and C. D. Shaw, Dynamics–the Geometry of Behavior , 2nd ed., Studies in Nonlinearity, Vol. Part Three: Global Behavior (Addison-Wesley, Advanced Book Program, Redwood City, Calif, 1992)
work page 1992
-
[38]
F. J. Varela, Principles of Biological Autonomy , new annotated edition ed., edited by E. A. Di Paolo and E. Thompson (The MIT Press, Cambridge, Mas- sachusetts London, 2025)
work page 2025
-
[39]
H. R. Maturana, F. J. Varela, S. Beer, and H. R. Mat- urana, Autopoiesis and Cognition: The Realization of the Living , Boston Studies in the Philosophy of Sci- ence No. 42 (D. Reidel Publishing Company, Dordrecht Boston London, 1980)
work page 1980
-
[40]
R. D. Beer and P. L. Williams, Animals and Animats: Why Not Both Iguanas?, Adaptive Behavior 17, 296 (2009)
work page 2009
-
[41]
R. D. Beer, A.-S. Barwich, and G. J. Severino, Milking a spherical cow: Toy models in neuroscience, Eur J of Neuroscience 60, 6359 (2024)
work page 2024
-
[42]
S. A. Kauffman, Investigations (Oxford University Press- New York, NY, 2000)
work page 2000
-
[43]
Alon, An Introduction to Systems Biology: Design Principles of Biological Circuits , 2nd ed
U. Alon, An Introduction to Systems Biology: Design Principles of Biological Circuits , 2nd ed. (Chapman and Hall/CRC, Second edition. | Boca Raton, Fla. : CRC Press, [2019], 2019)
work page 2019
-
[44]
C. McShaffrey and R. D. Beer, Decomposing Viability Space, in The 2023 Conference on Artificial Life (MIT Press, 2023)
work page 2023
-
[45]
Y. Kalinin, S. Neumann, V. Sourjik, and M. Wu, Re- sponses of Escherichia coli Bacteria to Two Opposing Chemoattractant Gradients Depend on the Chemorecep- tor Ratio, J Bacteriol 192, 1796 (2010)
work page 2010
-
[46]
C. McShaffrey and R. D. Beer, Dissecting Viability in Multi-Agent Systems, in The 2024 Conference on Artifi- cial Life (MIT Press, Online, 2024)
work page 2024
-
[47]
J. R. Collier, N. A. Monk, P. K. Maini, and J. H. Lewis, Pattern Formation by Lateral Inhibition with Feedback: A Mathematical Model of Delta-Notch Intercellular Sig- nalling, Journal of Theoretical Biology 183, 429 (1996)
work page 1996
-
[48]
J. S. Lowengrub, H. B. Frieboes, F. Jin, Y.-L. Chuang, X. Li, P. Macklin, S. M. Wise, and V. Cristini, Nonlinear modelling of cancer: Bridging the gap between cells and tumours, Nonlinearity 23, R1 (2010)
work page 2010
-
[49]
J. E. Ferrell, Bistability, Bifurcations, and Waddington’s Epigenetic Landscape, Current Biology 22, R458 (2012)
work page 2012
-
[50]
A. Raju and E. D. Siggia, A geometrical perspective on development, Dev Growth Differ 65, 245 (2023)
work page 2023
-
[51]
Y. Goyal, G. T. Busch, M. Pillai, J. Li, R. H. Boe, E. I. Grody, M. Chelvanambi, I. P. Dardani, B. Emert, N. Bodkin, J. Braun, D. Fingerman, A. Kaur, N. Jain, P. T. Ravindran, I. A. Mellis, K. Kiani, G. M. Alicea, M. E. Fane, S. S. Ahmed, H. Li, Y. Chen, C. Chai, J. Kaster, R. G. Witt, R. Lazcano, D. R. Ingram, S. B. Johnson, K. Wani, M. C. Dunagin, A. J....
work page 2023
- [52]
-
[53]
X. Qiu, Y. Zhang, J. D. Martin-Rufino, C. Weng, S. Hos- seinzadeh, D. Yang, A. N. Pogson, M. Y. Hein, K. Hoi (Joseph) Min, L. Wang, E. I. Grody, M. J. Shurtleff, R. Yuan, S. Xu, Y. Ma, J. M. Replogle, E. S. Lander, S. Darmanis, I. Bahar, V. G. Sankaran, J. Xing, and J. S. Weissman, Mapping transcriptomic vector fields of single cells, Cell 185, 690 (2022). 24
work page 2022
- [54]
-
[55]
S. Islam and S. Bhattacharya, Dynamical systems theory as an organizing principle for single-cell biology, npj Syst Biol Appl 11, 85 (2025)
work page 2025
-
[56]
Alon, Systems Medicine: Physiological Circuits and the Dynamics of Disease , 1st ed
U. Alon, Systems Medicine: Physiological Circuits and the Dynamics of Disease , 1st ed. (Chapman and Hall/CRC, Boca Raton, 2023)
work page 2023
-
[57]
E. Forbes and C. McShaffrey, Analyzing minimum viable populations in deterministic community models using vi- ability space decomposition (2026)
work page 2026
-
[58]
J. Aguirre, R. L. Viana, and M. A. F. Sanjuán, Fractal structures in nonlinear dynamics, Rev. Mod. Phys. 81, 333 (2009)
work page 2009
-
[59]
B. Krauskopf, H. M. Osinga, E. J. Doedel, M. E. Hender- son, J. Guckenheimer, A. Vladimirsky, M. Dellnitz, and O. Junge, A survey of methods for computing (un)stable manifolds of vector fields, Int. J. Bifurcation Chaos 15, 763 (2005)
work page 2005
-
[60]
C. McShaffrey and R. D. Beer, Shaking up viability space: Stochasticity and survival in the transient, in The 2025 Conference on Artificial Life (MIT Press, 2025)
work page 2025
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.