Fixation location in structured populations
Pith reviewed 2026-06-30 11:21 UTC · model grok-4.3
The pith
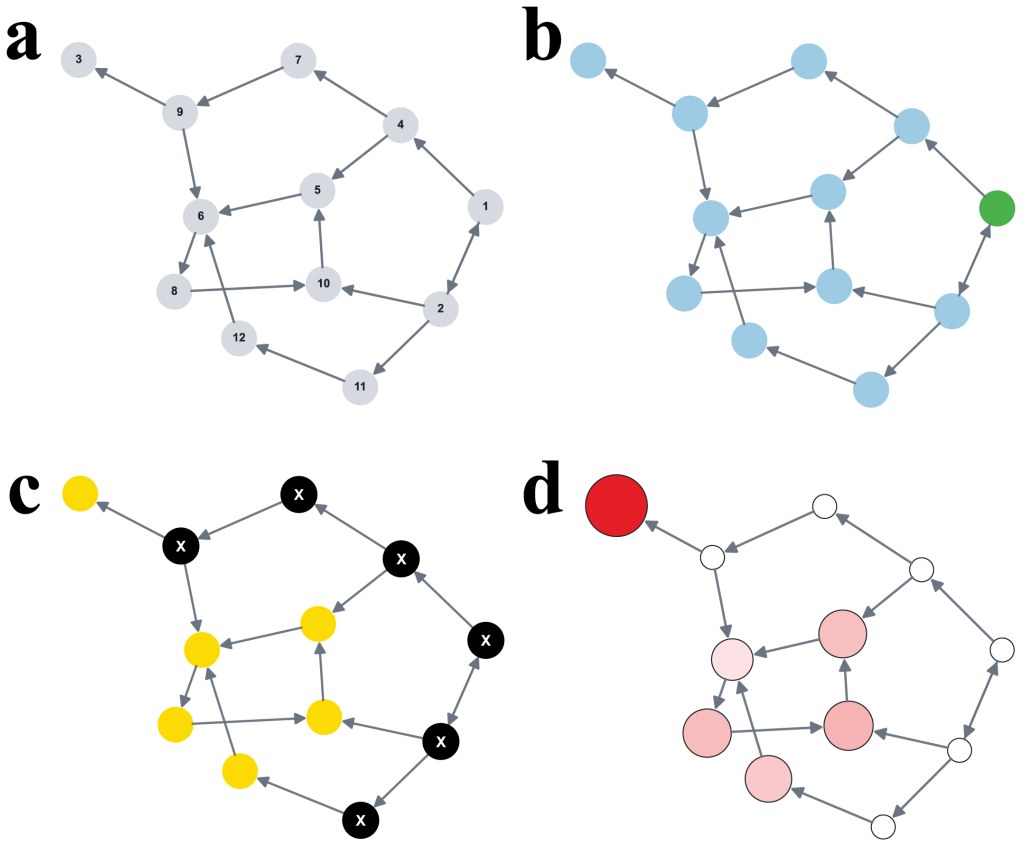
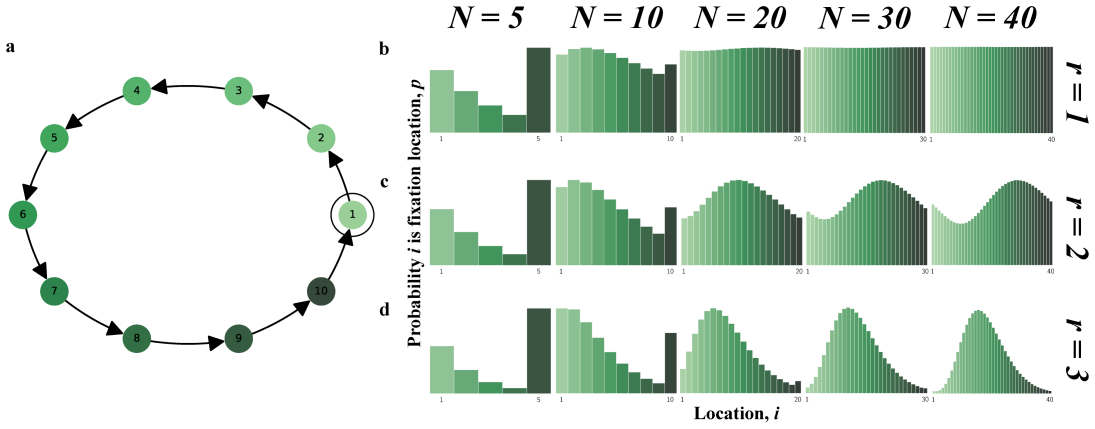
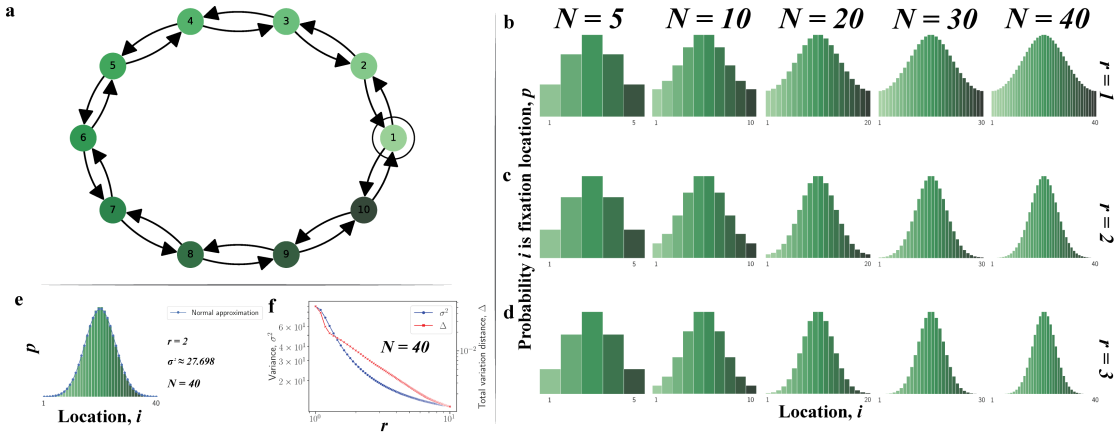
The node hosting the last wild-type before mutant fixation follows a nonuniform probability distribution set by graph structure and selection strength.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
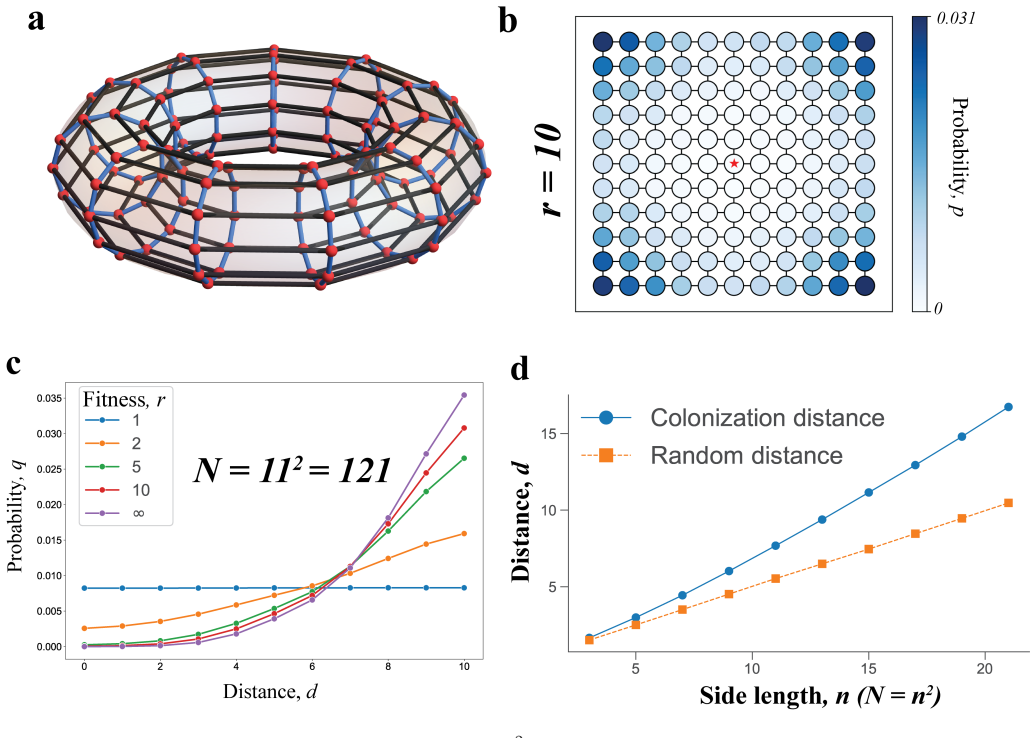
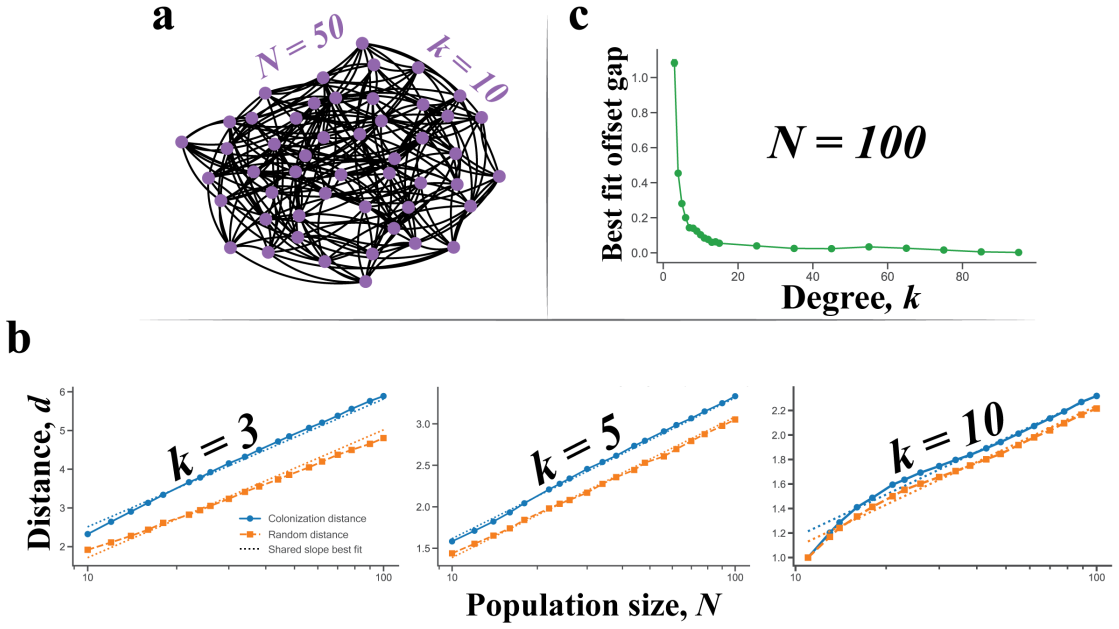
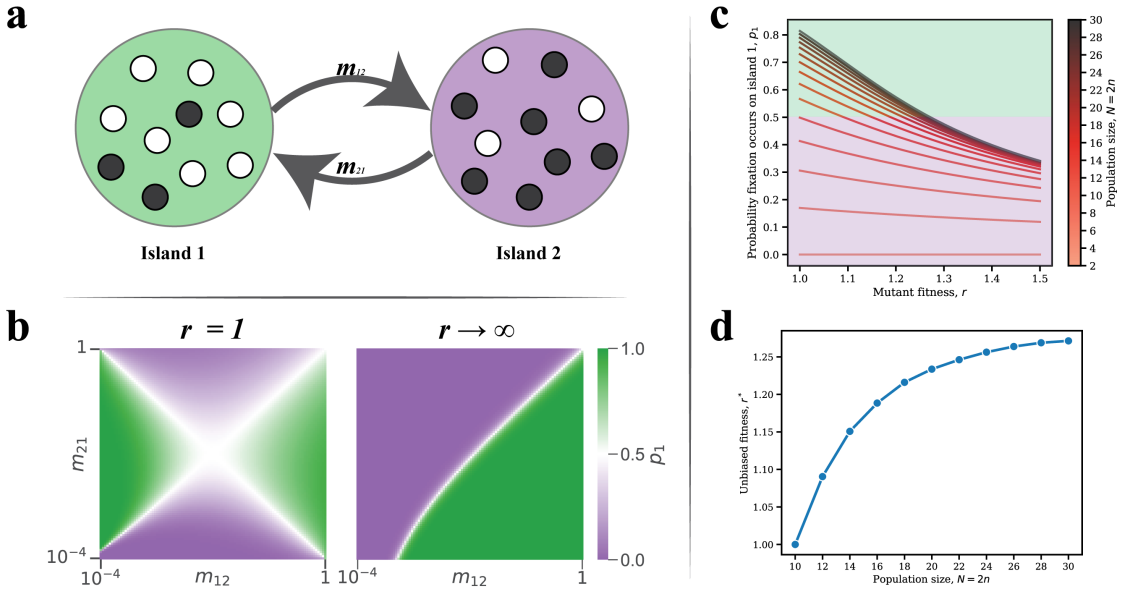
Conditional on fixation, the fixation location is the node occupied by the last wild-type individual immediately before mutant fixation. This location is described by a probability distribution over the nodes of the graph. The distribution is often highly nonuniform, depends strongly on the graph structure and the selection strength, and can differ sharply even when classical fixation statistics are similar. For many graphs some nodes can never be fixation locations. The result holds for neutral evolution, colonization, and constant selection on small graphs, cycles, tori, random graphs, and island populations.
What carries the argument
Conditional probability distribution of the fixation location, obtained from the transition probabilities of the Markov chain describing birth-death updating on the graph.
If this is right
- The fixation-location distribution supplies information about evolutionary outcomes that is not contained in fixation probability or mean fixation time.
- Certain nodes on many graphs are impossible sites for fixation, so the process cannot eliminate the wild-type last at those positions.
- The nonuniformity changes with selection strength, becoming more or less pronounced as selection moves from neutral to strong.
- The same qualitative features appear on cycles, tori, random graphs, and island models, indicating the phenomenon is not limited to special topologies.
Where Pith is reading between the lines
- Network designers could select topologies that make the fixation location more predictable or that protect particular nodes from serving as the final replacement site.
- In cultural or opinion dynamics the fixation location would correspond to the last individual holding an old trait, offering a spatial marker for tracking large-scale change.
- Empirical tracking of the final wild-type position in spatially structured microbial or tumor populations could test the predicted distributions directly.
- The same conditional-distribution calculation could be repeated for death-birth updating or frequency-dependent fitness to check whether nonuniformity persists.
Load-bearing premise
The evolutionary process on the graph is a Markovian birth-death updating rule under neutral evolution, colonization, or constant selection that permits the conditional distribution of the last wild-type node to be computed from the transition probabilities.
What would settle it
Simulations or exact calculations on a cycle graph under neutral drift showing that every node has exactly the same probability of being the fixation location, or a node with theoretically zero probability appearing as the fixation location in any realization.
Figures
read the original abstract
In stochastic evolutionary dynamics, the replacement of an existing genotype or cultural trait by a newly introduced mutant is typically characterized by the quantities of fixation probability and fixation time. But in a structured population, the disappearance of a lineage occurs at a specific place. For evolutionary dynamics on graphs, we define the fixation location as the node occupied by the last wild-type individual immediately before mutant fixation. Conditional on fixation, this location is described by a probability distribution over the nodes of the graph. We study the fixation location for neutral evolution, for the colonization process, and, more generally, for constant selection on small graphs, cycles, tori, random graphs, and island populations. We find that the distribution of the fixation location is often highly nonuniform, depends strongly on the graph structure and the selection strength, and can differ sharply even when classical fixation statistics are similar. For many graphs, some nodes can never be fixation locations. Our results identify fixation location as a fundamental aspect of evolutionary dynamics and suggest new ways to understand, monitor, and potentially mitigate extinction events in biological and social settings.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript introduces the fixation location as the node occupied by the last wild-type individual immediately before mutant fixation in birth-death processes on graphs. Conditional on fixation, it defines and computes the probability distribution of this location under neutral evolution, colonization, and constant selection. The authors examine this distribution on small graphs, cycles, tori, random graphs, and island models, reporting that it is frequently highly nonuniform, depends on graph structure and selection strength, can differ markedly even when fixation probability and time are similar, and that certain nodes are unreachable as fixation locations.
Significance. If the reported distributions hold, the work adds a spatially resolved observable to evolutionary graph theory that is not reducible to fixation probability or time. The systematic comparison across neutral, colonization, and selection regimes on standard graph families is a strength, as is the observation that some nodes are structurally excluded from being fixation locations. This could inform models of local extinction in biological and social networks.
minor comments (3)
- The manuscript should explicitly state the precise birth-death update rule (e.g., whether birth or death is selected first) and the normalization of fitness-dependent probabilities in the methods section, as these choices affect the transition matrix used to compute the conditional distribution.
- Figure captions and axis labels for the reported distributions should include the exact parameter values (selection strength, graph size) used in each panel to allow direct reproduction.
- A brief discussion of computational method (exact solution of the absorbing Markov chain versus Monte Carlo sampling) and any truncation or approximation employed for larger graphs would strengthen the results section.
Simulated Author's Rebuttal
We thank the referee for the supportive summary and recommendation of minor revision. The assessment correctly identifies the core contribution: that the fixation location distribution is a distinct, spatially resolved observable that is not reducible to fixation probability or time and that varies with graph structure and selection regime. No major comments were provided in the report.
Circularity Check
No significant circularity
full rationale
The paper introduces fixation location as a new observable (the node of the final wild-type individual before absorption) and derives its conditional distribution directly from the one-step transition matrix of the birth-death Markov chain on the graph. This computation is a standard consequence of solving the absorbing chain for the probability of each possible last-wild-type state; it requires no fitted parameters, no self-referential equations, and no load-bearing self-citations. The reported nonuniformity and impossible nodes are immediate algebraic outcomes of the adjacency matrix and fitness-dependent birth rates, with no reduction of the claimed results to the inputs by construction.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Evolutionary replacement on graphs follows a stochastic birth-death process determined by fitness and graph edges.
Reference graph
Works this paper leans on
-
[1]
Xxi.—on the dominance ratio
Ronald A Fisher. Xxi.—on the dominance ratio. Proceedings of the royal society of Edinburgh , 42:321--341, 1923
1923
-
[2]
A mathematical theory of natural and artificial selection, part v: selection and mutation
John Burdon Sanderson Haldane. A mathematical theory of natural and artificial selection, part v: selection and mutation. In Mathematical proceedings of the Cambridge philosophical society , volume 23, pages 838--844. Cambridge University Press, 1927
1927
-
[3]
Evolution in mendelian populations
Sewall Wright. Evolution in mendelian populations. Genetics , 16(2):97–159, March 1931
1931
-
[4]
R. A. Fisher and E. B. Ford. The "sewall wright effect. Heredity , 4:117–19, 1950
1950
-
[5]
Random processes in genetics
Patrick Alfred Pierce Moran. Random processes in genetics. In Mathematical proceedings of the cambridge philosophical society , volume 54, pages 60--71. Cambridge University Press, 1958
1958
-
[6]
On the probability of fixation of mutant genes in a population
Motoo Kimura. On the probability of fixation of mutant genes in a population. Genetics , 47:713--719, 1962
1962
-
[7]
The genetical evolution of social behaviour
William D Hamilton. The genetical evolution of social behaviour. ii. Journal of theoretical biology , 7(1):17--52, 1964
1964
-
[8]
Cultural transmission and evolution: A quantitative approach
Luigi Luca Cavalli-Sforza and Marcus W Feldman. Cultural transmission and evolution: A quantitative approach . Princeton University Press, 1981
1981
-
[9]
Culture and the evolutionary process
Robert Boyd and Peter J Richerson. Culture and the evolutionary process . University of Chicago press, 1988
1988
-
[10]
Evolution in Age-Structured Populations
Brian Charlesworth. Evolution in Age-Structured Populations . Cambridge University Press, Cambridge, 2 edition, 1994
1994
-
[11]
Mathematical population genetics: theoretical introduction , volume 27
Warren John Ewens. Mathematical population genetics: theoretical introduction , volume 27. Springer, 2004
2004
-
[12]
Five rules for the evolution of cooperation
Martin A Nowak. Five rules for the evolution of cooperation. Science , 314(5805):1560--1563, 2006
2006
-
[13]
Patrick A. P. Moran. The Statistical Processes of Evolutionary Theory . Oxford University Press, 1962
1962
-
[14]
Nowak, Akira Sasaki, Christine Taylor, and Drew Fudenberg
Martin A. Nowak, Akira Sasaki, Christine Taylor, and Drew Fudenberg. Emergence of cooperation and evolutionary stability in finite populations. Nature , 428:646--650, 2004
2004
-
[15]
Evolutionary dynamics: exploring the equations of life
Martin A Nowak. Evolutionary dynamics: exploring the equations of life . Harvard University Press, 2006
2006
-
[16]
Evolutionary dynamics on graphs
Erez Lieberman, Christoph Hauert, and Martin A Nowak. Evolutionary dynamics on graphs. Nature , 433(7023):312--316, 2005
2005
-
[17]
Evolutionary games on cycles
Hisashi Ohtsuki and Martin A Nowak. Evolutionary games on cycles. Proceedings of the Royal Society B: Biological Sciences , 273(1598):2249--2256, 2006
2006
-
[18]
Evolutionary graph theory: Breaking the symmetry between interaction and replacement
Hisashi Ohtsuki, Jorge M Pacheco, and Martin A Nowak. Evolutionary graph theory: Breaking the symmetry between interaction and replacement. Journal of Theoretical Biology , 246(4):681--694, 2007
2007
-
[19]
Evolutionary stability on graphs
Hisashi Ohtsuki and Martin A Nowak. Evolutionary stability on graphs. Journal of Theoretical Biology , 251(4):698--707, 2008
2008
-
[20]
Categorizing update mechanisms for graph-structured metapopulations
Sedigheh Yagoobi, Nikhil Sharma, and Arne Traulsen. Categorizing update mechanisms for graph-structured metapopulations. Journal of the Royal Society Interface , 20(200), 2023
2023
-
[21]
Most undirected random graphs are amplifiers of selection for birth-death dynamics, but suppressors of selection for death-birth dynamics
Laura Hindersin and Arne Traulsen. Most undirected random graphs are amplifiers of selection for birth-death dynamics, but suppressors of selection for death-birth dynamics. PLoS computational biology , 11(11):e1004437, 2015
2015
-
[22]
Andreas Pavlogiannis, Josef Tkadlec, Krishnendu Chatterjee, and Martin A. Nowak. Amplification on undirected population structures: Comets beat stars. Scientific Reports , 7:82, 2017
2017
-
[23]
Andreas Pavlogiannis, Josef Tkadlec, Krishnendu Chatterjee, and Martin A. Nowak. Construction of arbitrarily strong amplifiers of natural selection using evolutionary graph theory. Communications Biology , 1:71, 2018
2018
-
[24]
Transient amplifiers of selection and reducers of fixation for death-birth updating on graphs
Benjamin Allen et al. Transient amplifiers of selection and reducers of fixation for death-birth updating on graphs. PLOS Computational Biology , 16:1--20, 2020
2020
-
[25]
Limits on amplifiers of natural selection under death-birth updating
Josef Tkadlec, Andreas Pavlogiannis, Krishnendu Chatterjee, and Martin A Nowak. Limits on amplifiers of natural selection under death-birth updating. PLoS computational biology , 16(1):e1007494, 2020
2020
-
[26]
Suppressors of fixation can increase average fitness beyond amplifiers of selection
Nikhil Sharma and Arne Traulsen. Suppressors of fixation can increase average fitness beyond amplifiers of selection. Proceedings of the National Academy of Sciences , 119(37):e2205424119, 2022
2022
-
[27]
Self-loops in evolutionary graph theory: Friends or foes? PLoS Computational Biology , 19(9):e1011387, 2023
Nikhil Sharma, Sedigheh Yagoobi, and Arne Traulsen. Self-loops in evolutionary graph theory: Friends or foes? PLoS Computational Biology , 19(9):e1011387, 2023
2023
-
[28]
Graph-structured populations elucidate the role of deleterious mutations in long-term evolution
Nikhil Sharma, Suman G Das, Joachim Krug, and Arne Traulsen. Graph-structured populations elucidate the role of deleterious mutations in long-term evolution. Nature Communications , 16(1):2355, 2025
2025
-
[29]
The effect of population structure on the rate of evolution
Marcus Frean, Paul B Rainey, and Arne Traulsen. The effect of population structure on the rate of evolution. Proceedings of the Royal Society B: Biological Sciences , 280(1762), 2013
2013
-
[30]
The molecular clock of neutral evolution can be accelerated or slowed by asymmetric spatial structure
Benjamin Allen, Christine Sample, Yulia Dementieva, Ruben C Medeiros, Christopher Paoletti, and Martin A Nowak. The molecular clock of neutral evolution can be accelerated or slowed by asymmetric spatial structure. PLoS Computational Biology , 11(2):e1004108, 2015
2015
-
[31]
Population structure determines the tradeoff between fixation probability and fixation time
Josef Tkadlec, Andreas Pavlogiannis, Krishnendu Chatterjee, and Martin A Nowak. Population structure determines the tradeoff between fixation probability and fixation time. Communications biology , 2(1):138, 2019
2019
-
[32]
Maintaining diversity in structured populations
David A Brewster, Jakub Svoboda, Dylan Roscow, Krishnendu Chatterjee, Josef Tkadlec, and Martin A Nowak. Maintaining diversity in structured populations. PNAS nexus , 4(8):pgaf252, 2025
2025
-
[33]
Population structure amplifies mutation load
Nikhil Sharma, Suman G Das, Arne Traulsen, and Joachim Krug. Population structure amplifies mutation load. bioRxiv , pages 2025--09, 2025
2025
-
[34]
Nowak and Robert M
Martin A. Nowak and Robert M. May. Evolutionary games and spatial chaos. Nature , 359(63986398):826–829, October 1992
1992
-
[35]
Evolutionary dynamics on any population structure
Benjamin Allen, Gabor Lippner, Yu-Ting Chen, Babak Fotouhi, Naghmeh Momeni, Shing-Tung Yau, and Martin A Nowak. Evolutionary dynamics on any population structure. Nature , 544(7649):227--230, 2017
2017
-
[36]
An analysis of the fixation probability of a mutant on special classes of non-directed graphs
Mark Broom and Jan Rycht \'a r . An analysis of the fixation probability of a mutant on special classes of non-directed graphs. Proceedings of the Royal Society A: Mathematical, Physical and Engineering Sciences , 464:2609--2627, 2008
2008
-
[37]
Two results on evolutionary processes on general non-directed graphs
M Broom, C Hadjichrysanthou, J Rycht \'a r , and BT Stadler. Two results on evolutionary processes on general non-directed graphs. Proceedings of the Royal Society A: Mathematical, Physical and Engineering Sciences , 466(2121):2795--2798, 2010
2010
-
[38]
Mark Broom, Jan Rycht \'a r , and Boris T. Stadler. Evolutionary dynamics on graphs - the effect of graph structure and initial placement on mutant spread. Journal of Statistical Theory and Practice , 5:369--381, 2011
2011
-
[39]
The duality of spatial death--birth and birth--death processes and limitations of the isothermal theorem
Kamran Kaveh, Natalia L Komarova, and Mohammad Kohandel. The duality of spatial death--birth and birth--death processes and limitations of the isothermal theorem. Royal Society open science , 2(4):140465, 2015
2015
-
[40]
Spatial structure facilitates evolutionary rescue by drug resistance
Cecilia Fruet, Ella Linxia M \"u ller, Claude Loverdo, and Anne-Florence Bitbol. Spatial structure facilitates evolutionary rescue by drug resistance. PLOS Computational Biology , 21(4):e1012861, 2025
2025
-
[41]
Mixed updating in structured populations
David A Brewster, Yichen Huang, Michael Mitzenmacher, and Martin A Nowak. Mixed updating in structured populations. arXiv preprint arXiv:2512.11164 , 2025
-
[42]
Evolutionary games on graphs and the speed of the evolutionary process
Mark Broom, Christophoros Hadjichrysanthou, and Jan Rycht \'a r . Evolutionary games on graphs and the speed of the evolutionary process. Proceedings of the Royal Society A: Mathematical, Physical and Engineering Sciences , 466(2117):1327--1346, 2010
2010
-
[43]
Approximating fixation probabilities in the generalized moran process
Josep D \' az, Leslie Ann Goldberg, George B Mertzios, David Richerby, Maria Serna, and Paul G Spirakis. Approximating fixation probabilities in the generalized moran process. Algorithmica , 69:78--91, 2014
2014
-
[44]
Absorption time of the moran process
Josep D \' az, Leslie Ann Goldberg, David Richerby, and Maria Serna. Absorption time of the moran process. Random Structures & Algorithms , 49(1):137--159, 2016
2016
-
[45]
Fitness dependence of the fixation-time distribution for evolutionary dynamics on graphs
David Hathcock and Steven H Strogatz. Fitness dependence of the fixation-time distribution for evolutionary dynamics on graphs. Physical Review E , 100(1):012408, 2019
2019
-
[46]
Wald’s martingale and the conditional distributions of absorption time in the moran process
Travis Monk and Andr \'e van Schaik. Wald’s martingale and the conditional distributions of absorption time in the moran process. Proceedings of the Royal Society A , 476(2241):20200135, 2020
2020
-
[47]
Evolutionary graph theory beyond pairwise interactions: higher-order network motifs shape times to fixation in structured populations
Yang Ping Kuo and Oana Carja. Evolutionary graph theory beyond pairwise interactions: higher-order network motifs shape times to fixation in structured populations. PLOS Computational Biology , 20(3):e1011905, 2024
2024
-
[48]
Fixation times on directed graphs
David A Brewster, Martin A Nowak, and Josef Tkadlec. Fixation times on directed graphs. PLOS Computational Biology , 20(7):e1012299, 2024
2024
-
[49]
Colonization times in moran process on graphs
Lenka Kopfov \'a and Josef Tkadlec. Colonization times in moran process on graphs. PLoS computational biology , 21(5):e1012868, 2025
2025
-
[50]
Random walk: a modern introduction , volume 123
Gregory F Lawler and Vlada Limic. Random walk: a modern introduction , volume 123. Cambridge University Press, 2010
2010
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.