Compositional proofreading through critical self-tuning
Pith reviewed 2026-06-27 10:39 UTC · model grok-4.3
The pith
Competition for shared inputs pins multicomponent systems to the marginal stability threshold of the strongest species, implementing natural proofreading.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
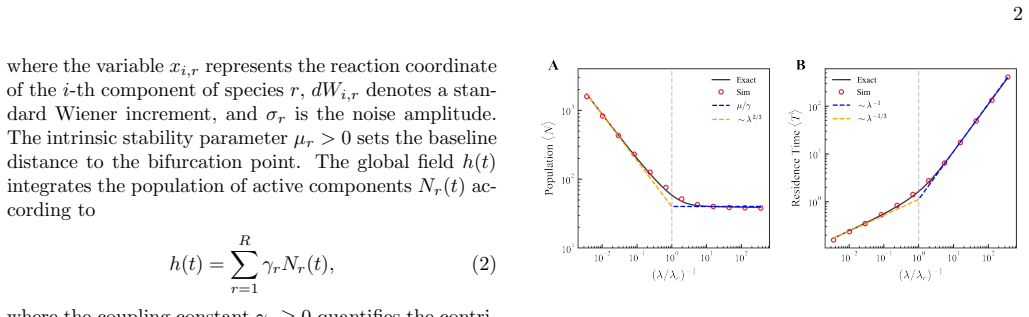
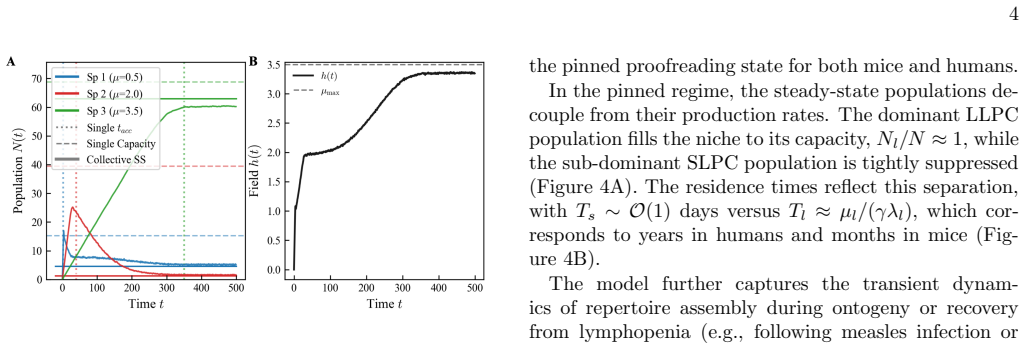
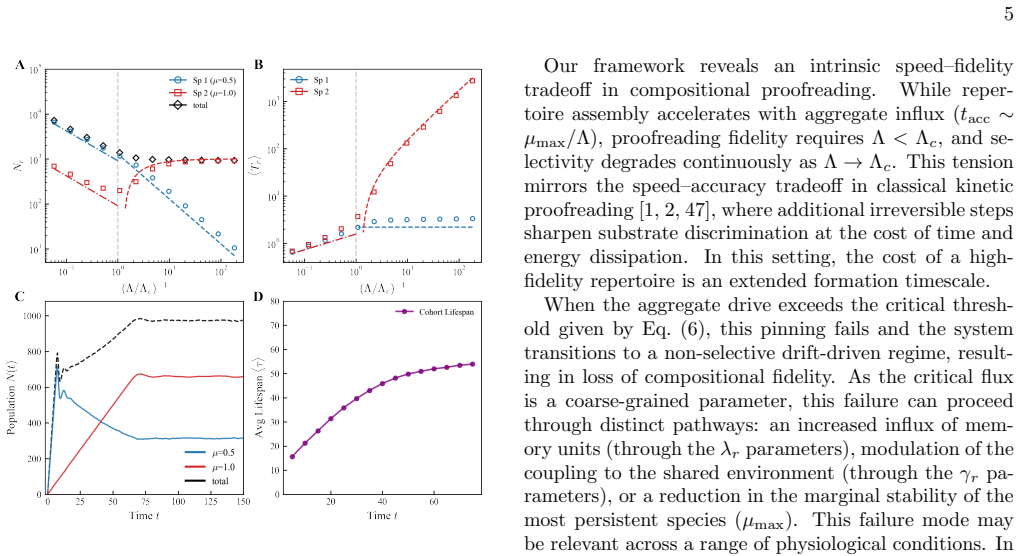
The paper claims that competition for shared inputs naturally pins the system to the marginal stability threshold of the most persistent species. This grants dominant species extended lifetimes, concentrating the population into dominant components while forcing less-stable variants into rapid drift-driven turnover, thereby implementing compositional proofreading. When aggregate drive exceeds a characteristic scale, this pinning fails, producing a non-selective state where component lifetimes scale as a universal power law with aggregate drive. The same mechanism is proposed to explain hallmarks in plasma cell accumulation dynamics, with de-pinning transitions marking failure points in biolo
What carries the argument
Competition for shared inputs that pins the system to the marginal stability threshold of the most persistent species, thereby implementing compositional proofreading.
If this is right
- Dominant species receive extended lifetimes from the pinning effect.
- Less-stable variants undergo rapid drift-driven turnover.
- Above a characteristic aggregate drive the system loses selectivity and component lifetimes follow a universal power law.
- Signatures of the pinning appear in plasma cell accumulation dynamics.
- De-pinning transitions mark failure points in domains such as cancer, immunodeficiencies, and ageing-related genomic activation.
Where Pith is reading between the lines
- The same competition-driven pinning may operate in epigenetic repertoires without separate regulatory layers.
- Varying the strength of shared-input competition in cell-culture models could locate the de-pinning transition experimentally.
- Power-law lifetime scaling above the drive threshold offers a testable population-level signature in longitudinal biological data.
- Failure of the pinning mechanism could contribute to accumulation of suboptimal or harmful variants across multiple high-dimensional biological systems.
Load-bearing premise
Competition for shared inputs will spontaneously pin the system to the marginal stability threshold of the most persistent species without requiring additional tuning mechanisms or specific initial conditions beyond the competition itself.
What would settle it
Measure lifetimes of components in a competitive multicomponent system while varying aggregate drive; check whether the most persistent species show extended lifetimes below a threshold scale and whether all lifetimes shift to a common power-law scaling above that scale.
Figures

read the original abstract
High-dimensional multicomponent systems, including immune and epigenetic repertoires, must selectively retain rare, beneficial components while purging a massive influx of suboptimal variants. We demonstrate that critical tuning of component control parameters through competition naturally implements proofreading in these systems. Competition for shared inputs pins the system to the marginal stability threshold of the most persistent species. This grants dominant species extended lifetimes, concentrating the population into dominant components while forcing less-stable variants into rapid drift-driven turnover. When aggregate drive exceeds a characteristic scale, this pinning fails, producing a non-selective state where component lifetimes scale as a universal power law with aggregate drive. Applying this framework to biological memory, we identify the hallmarks of this effect in plasma cell accumulation dynamics and propose that de-pinning transitions may represent failure points across biological domains, including cancer, immunodeficiencies, and the aberrant activation of harmful genomic elements during ageing.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper claims that high-dimensional multicomponent systems achieve compositional proofreading via critical self-tuning: competition for shared inputs spontaneously pins the dynamics to the marginal stability threshold of the most persistent species, extending its lifetime while driving rapid turnover of less-stable variants. When aggregate drive exceeds a characteristic scale, pinning fails and lifetimes follow a universal power law. The framework is applied to plasma-cell accumulation dynamics, with de-pinning transitions proposed as failure modes in cancer, immunodeficiencies, and ageing.
Significance. If the pinning mechanism is shown to arise generically from competition without extra tuning, the work would supply a concrete, testable route to selectivity in repertoires that must retain rare beneficial components amid massive variant influx. The reported power-law regime supplies a falsifiable signature, and the plasma-cell application offers an immediate biological test case.
major comments (2)
- [Abstract] Abstract: the central assertion that 'competition for shared inputs pins the system to the marginal stability threshold of the most persistent species' is stated without model equations, stability analysis, or simulation protocol. Because this pinning is the load-bearing step that converts competition into proofreading, its spontaneous occurrence for generic initial conditions must be derived explicitly.
- [Application to plasma cell dynamics] Application section: the claim to 'identify the hallmarks of this effect in plasma cell accumulation dynamics' is made without reference to any dataset, figure, or quantitative comparison. This leaves the biological mapping unsupported and prevents assessment of whether the predicted lifetime scaling or de-pinning transition is actually observed.
minor comments (1)
- [Abstract] The phrase 'characteristic scale of aggregate drive' is introduced as the point at which pinning fails, yet no definition or derivation of this scale is supplied in the abstract; its status as a free parameter versus an emergent quantity should be clarified.
Simulated Author's Rebuttal
We thank the referee for the constructive comments, which help clarify the presentation of the core mechanism and its biological application. We address each point below.
read point-by-point responses
-
Referee: [Abstract] Abstract: the central assertion that 'competition for shared inputs pins the system to the marginal stability threshold of the most persistent species' is stated without model equations, stability analysis, or simulation protocol. Because this pinning is the load-bearing step that converts competition into proofreading, its spontaneous occurrence for generic initial conditions must be derived explicitly.
Authors: The full manuscript derives the pinning explicitly: Section 2 presents the model equations for the multicomponent system with shared inputs; Section 3 contains the linear stability analysis around the marginal stability threshold, proving that competition drives the system to the threshold of the most persistent species for generic initial conditions; Section 4 details the simulation protocol and shows the spontaneous pinning in numerical trajectories. The abstract summarizes this result concisely. We have revised the abstract to include a brief clause referencing the derivation and the generic initial-condition result. revision: yes
-
Referee: [Application to plasma cell dynamics] Application section: the claim to 'identify the hallmarks of this effect in plasma cell accumulation dynamics' is made without reference to any dataset, figure, or quantitative comparison. This leaves the biological mapping unsupported and prevents assessment of whether the predicted lifetime scaling or de-pinning transition is actually observed.
Authors: We agree the mapping requires stronger support. The original text identifies qualitative hallmarks (accumulation kinetics and turnover rates) drawn from existing literature on plasma-cell dynamics. In revision we add explicit citations to published datasets on plasma-cell lifetimes, include a new figure overlaying the model's predicted power-law scaling against reported empirical values, and discuss the de-pinning threshold in the context of observed failure modes. This supplies the quantitative comparison requested. revision: yes
Circularity Check
No significant circularity identified
full rationale
The paper's claimed mechanism—that competition for shared inputs pins the system to the marginal stability threshold of the most persistent species, granting extended lifetimes to dominant components—follows as a dynamical consequence of the stated competition rules rather than reducing to a self-definition, a fitted parameter renamed as a prediction, or a load-bearing self-citation. No equations or steps in the abstract or described framework exhibit the specific reductions required for circularity (e.g., an outcome defined in terms of itself or a uniqueness theorem imported solely from prior author work). The derivation remains self-contained against external benchmarks of model behavior.
Axiom & Free-Parameter Ledger
free parameters (1)
- characteristic scale of aggregate drive
axioms (1)
- domain assumption High-dimensional multicomponent systems must selectively retain rare beneficial components while purging a massive influx of suboptimal variants.
Reference graph
Works this paper leans on
-
[1]
J. J. Hopfield, Kinetic proofreading: a new mechanism for reducing errors in biosynthetic processes requiring high specificity, Proceedings of the National Academy of Sciences71, 4135 (1974)
1974
-
[2]
Ninio, Kinetic amplification of enzyme discrimination, Biochimie57, 587 (1975)
J. Ninio, Kinetic amplification of enzyme discrimination, Biochimie57, 587 (1975)
1975
-
[3]
T. W. McKeithan, Kinetic proofreading in t-cell re- ceptor signal transduction., Proceedings of the national academy of sciences92, 5042 (1995)
1995
-
[4]
Savir and T
Y. Savir and T. Tlusty, Conformational proofreading: the impact of conformational changes on the specificity of molecular recognition, PloS one2, e468 (2007)
2007
-
[5]
Murugan, D
A. Murugan, D. A. Huse, and S. Leibler, Discriminatory proofreading regimes in nonequilibrium systems, Physi- cal Review X4, 021016 (2014)
2014
-
[6]
R. Ravasio, K. Husain, C. G. Evans, R. Phillips, M. Ribezzi, J. W. Szostak, and A. Murugan, A minimal scenario for the origin of non-equilibrium order, arXiv preprint arXiv:2405.10911 (2024)
-
[7]
Tonegawa, Somatic generation of antibody diversity, Nature302, 575 (1983)
S. Tonegawa, Somatic generation of antibody diversity, Nature302, 575 (1983)
1983
-
[8]
Radbruch, G
A. Radbruch, G. Muehlinghaus, E. O. Luger, A. In- amine, K. G. Smith, T. D¨ orner, and F. Hiepe, Com- petence and competition: the challenge of becoming a long-lived plasma cell, Nature Reviews Immunology6, 741 (2006)
2006
-
[9]
Altan-Bonnet, T
G. Altan-Bonnet, T. Mora, and A. M. Walczak, Quanti- tative immunology for physicists, Physics Reports849, 1 (2020)
2020
-
[10]
B. D. Simons and O. Karin, Tuning of plasma cell lifespan by competition explains the longevity and heterogeneity of antibody persistence, Immunity57, 600 (2024)
2024
-
[11]
Beltran, V
T. Beltran, V. Shahrezaei, V. Katju, and P. Sarkies, Epimutations driven by small rnas arise frequently but most have limited duration in caenorhabditis elegans, Na- ture ecology & evolution4, 1539 (2020)
2020
-
[12]
Karin, E
O. Karin, E. A. Miska, and B. D. Simons, Epigenetic in- heritance of gene silencing is maintained by a self-tuning mechanism based on resource competition, Cell systems 14, 24 (2023)
2023
-
[13]
Purves and J
D. Purves and J. W. Lichtman, Elimination of synapses in the developing nervous system, Science210, 153 (1980)
1980
-
[14]
C. S. Goodman and C. J. Shatz, Developmental mecha- nisms that generate precise patterns of neuronal connec- tivity, Cell72, 77 (1993)
1993
-
[15]
Bentley, G
K. Bentley, G. Mariggi, H. Gerhardt, and P. A. Bates, Tipping the balance: robustness of tip cell selection, mi- gration and fusion in angiogenesis, PLoS computational biology5, e1000549 (2009)
2009
-
[16]
S. M. van Neerven and L. Vermeulen, Cell competition in development, homeostasis and cancer, Nature Reviews Molecular Cell Biology24, 221 (2023)
2023
-
[17]
B. D. Simons and O. Karin, Cell cycle criticality as a mechanism for robust cell population control, Molecular Systems Biology , 1 (2025)
2025
-
[18]
Gerhart and M
J. Gerhart and M. Kirschner, The theory of facilitated variation, Proceedings of the National Academy of Sci- ences104, 8582 (2007)
2007
-
[19]
Bagci, Y
E. Bagci, Y. Vodovotz, T. Billiar, G. Ermentrout, and I. Bahar, Bistability in apoptosis: roles of bax, bcl-2, and mitochondrial permeability transition pores, Biophysical journal90, 1546 (2006)
2006
-
[20]
J. G. Albeck, J. M. Burke, S. L. Spencer, D. A. Lauf- fenburger, and P. K. Sorger, Modeling a snap-action, variable-delay switch controlling extrinsic cell death, PLoS biology6, e299 (2008)
2008
-
[21]
G. Yao, C. Tan, M. West, J. R. Nevins, and L. You, Ori- gin of bistability underlying mammalian cell cycle entry, Molecular systems biology7, 485 (2011)
2011
-
[22]
Briffa, G
A. Briffa, G. Menon, A. Movilla Miangolarra, and M. Howard, Dissecting mechanisms of epigenetic mem- ory through computational modeling, Annual Review of Plant Biology75, 265 (2024)
2024
-
[23]
Stanoev, A
A. Stanoev, A. P. Nandan, and A. Koseska, Organization at criticality enables processing of time-varying signals by receptor networks, Molecular systems biology16, e8870 (2020)
2020
-
[24]
D. Koch, A. Nandan, G. Ramesan, and A. Koseska, Biological computations: limitations of attractor-based formalisms and the need for transients, Biochemical and Biophysical Research Communications720, 150069 (2024)
2024
-
[25]
Mora and W
T. Mora and W. Bialek, Are biological systems poised at criticality?, Journal of Statistical Physics144, 268 (2011)
2011
-
[26]
T. Agranov, N. Wiegenfeld, O. Karin, and B. D. Si- mons, Self-organized hyperuniformity in a minimal model of population dynamics, arXiv preprint arXiv:2509.08077 (2025)
-
[27]
D. S. Fisher, Collective transport in random media: from superconductors to earthquakes, Physics reports301, 113 (1998)
1998
-
[28]
Reichhardt and C
C. Reichhardt and C. J. Olson Reichhardt, Depinning and nonequilibrium dynamic phases of particle assem- blies driven over random and ordered substrates: a re- view, Reports on Progress in Physics80, 026501 (2017)
2017
-
[29]
Knebel, M
J. Knebel, M. F. Weber, T. Kr¨ uger, and E. Frey, Evo- lutionary games of condensates in coupled birth–death processes, Nature communications6, 6977 (2015)
2015
-
[30]
S. F. Brynjolfsson, L. Persson Berg, T. Olsen Eker- hult, I. Rimkute, M.-J. Wick, I.-L. M˚ artensson, and O. Grimsholm, Long-lived plasma cells in mice and men, Frontiers in immunology9, 2673 (2018)
2018
-
[31]
Z. Jing, P. Galbo, L. Ovando, M. Demouth, S. Welte, R. Park, K. Chandran, Y. Wu, T. MacCarthy, D. Zheng, et al., Fine-tuning spatial-temporal dynamics and sur- face receptor expression support plasma cell-intrinsic longevity, Elife12, RP89712 (2024)
2024
-
[32]
Benet, Z
Z. Benet, Z. Jing, and D. R. Fooksman, Plasma cell dy- namics in the bone marrow niche, Cell reports34(2021)
2021
-
[33]
Eslami, S
M. Eslami, S. Schuepbach-Mallepell, D. Diana, L. Willen, C. Kowalczyk-Quintas, C. Desponds, B. Peter, M. Vigolo, F. Renevey, O. Donz´ e,et al., Unique and redundant roles of mouse bcma, taci, baff, april, and il-6 in supporting antibody-producing cells in different tissues, Proceedings of the National Academy of Sciences121, e2404309121 (2024)
2024
-
[34]
R. Park, Z. Benet, Z. Jing, J. Enright, and D. R. Fooks- man, Cd138 and april regulate plasma cell survival, com- petition, and retention in the bone marrow niche, Cell reports44(2025)
2025
-
[35]
R. D. de Vries, S. McQuaid, G. van Amerongen, S. Y¨ uksel, R. J. Verburgh, A. D. Osterhaus, W. P. Duprex, and R. L. de Swart, Measles immune suppres- 7 sion: Lessons from the macaque model, PLoS Pathogens 8, e1002885 (2012)
2012
-
[36]
M. J. Mina, C. J. E. Metcalf, R. L. De Swart, A. Os- terhaus, and B. T. Grenfell, Long-term measles-induced immunomodulation increases overall childhood infectious disease mortality, Science348, 694 (2015)
2015
-
[37]
A. Q. Xu, R. R. Barbosa, and D. P. Calado, Genetic timestamping of plasma cells in vivo reveals tissue- specific homeostatic population turnover, Elife9, e59850 (2020)
2020
-
[38]
X. Liu, J. Yao, Y. Zhao, J. Wang, and H. Qi, Heteroge- neous plasma cells and long-lived subsets in response to immunization, autoantigen and microbiota, Nature im- munology23, 1564 (2022)
2022
-
[39]
Koike, K
T. Koike, K. Fujii, K. Kometani, N. S. Butler, K. Fu- nakoshi, S. Yari, J. Kikuta, M. Ishii, T. Kurosaki, and W. Ise, Progressive differentiation toward the long-lived plasma cell compartment in the bone marrow, Journal of Experimental Medicine220, e20221717 (2022)
2022
-
[40]
M. J. Robinson, Z. Ding, M. R. Dowling, D. L. Hill, R. H. Webster, C. McKenzie, C. Pitt, K. O’Donnell, J. Mul- der, E. Brodie,et al., Intrinsically determined turnover underlies broad heterogeneity in plasma-cell lifespan, Im- munity56, 1596 (2023)
2023
-
[41]
Biajoux, J
V. Biajoux, J. Natt, C. Freitas, N. Alouche, A. Sac- quin, P. Hemon, F. Gaudin, N. Fazilleau, M. Espeli, and K. Balabanian, Efficient plasma cell differentiation and trafficking require cxcr4 desensitization, Cell reports17, 193 (2016)
2016
-
[42]
A. V. Gulino, D. Moratto, S. Sozzani, P. Cavadini, K. Otero, L. Tassone, L. Imberti, S. Pirovano, L. D. No- tarangelo, R. Soresina,et al., Altered leukocyte response to cxcl12 in patients with warts hypogammaglobuline- mia, infections, myelokathexis (whim) syndrome, Blood 104, 444 (2004)
2004
-
[43]
P. J. Mc Guire, C. Cunningham-Rundles, H. Ochs, and G. A. Diaz, Oligoclonality, impaired class switch and b- cell memory responses in whim syndrome, Clinical im- munology135, 412 (2010)
2010
-
[44]
Y.-T. Tai, C. Acharya, G. An, M. Moschetta, M. Y. Zhong, X. Feng, M. Cea, A. Cagnetta, K. Wen, H. van Eenennaam,et al., April and bcma promote human mul- tiple myeloma growth and immunosuppression in the bone marrow microenvironment, Blood, The Journal of the American Society of Hematology127, 3225 (2016)
2016
-
[45]
Chahin, Z
M. Chahin, Z. Branham, A. Fox, C. Leurinda, and A. R. Keruakous, Clinical considerations for immunoparesis in multiple myeloma, Cancers14, 2278 (2022)
2022
-
[46]
Karin, Learning principle for the adaptive shaping of epigenetic memory repertoires, PRX Life4, 023011 (2026)
O. Karin, Learning principle for the adaptive shaping of epigenetic memory repertoires, PRX Life4, 023011 (2026)
2026
-
[47]
Banerjee, A
K. Banerjee, A. B. Kolomeisky, and O. A. Igoshin, Eluci- dating interplay of speed and accuracy in biological error correction, Proceedings of the National Academy of Sci- ences114, 5183 (2017)
2017
-
[48]
J. G. Wood, B. C. Jones, N. Jiang, C. Chang, S. Hosier, P. Wickremesinghe, M. Garcia, D. A. Hartnett, L. Burhenn, N. Neretti,et al., Chromatin-modifying ge- netic interventions suppress age-associated transposable element activation and extend life span in drosophila, Proceedings of the National Academy of Sciences113, 11277 (2016)
2016
-
[49]
De Cecco, T
M. De Cecco, T. Ito, A. P. Petrashen, A. E. Elias, N. J. Skvir, S. W. Criscione, A. Caligiana, G. Brocculi, E. M. Adney, J. D. Boeke,et al., Line-1 derepression in senes- cent cells triggers interferon and inflammaging, Nature 566, 73 (2019)
2019
-
[50]
Lippman, A.-V
Z. Lippman, A.-V. Gendrel, M. Black, M. W. Vaughn, N. Dedhia, W. Richard McCombie, K. Lavine, V. Mit- tal, B. May, K. D. Kasschau,et al., Role of transposable elements in heterochromatin and epigenetic control, Na- ture430, 471 (2004)
2004
-
[51]
I. B. Dodd, M. A. Micheelsen, K. Sneppen, and G. Thon, Theoretical analysis of epigenetic cell memory by nucle- osome modification, Cell129, 813 (2007)
2007
-
[52]
Oberdoerffer, S
P. Oberdoerffer, S. Michan, M. McVay, R. Mostoslavsky, J. Vann, S.-K. Park, A. Hartlerode, J. Stegmuller, A. Hafner, P. Loerch,et al., Sirt1 redistribution on chro- matin promotes genomic stability but alters gene expres- sion during aging, Cell135, 907 (2008)
2008
-
[53]
Seczynska, S
M. Seczynska, S. Bloor, S. M. Cuesta, and P. J. Lehner, Genome surveillance by hush-mediated silencing of in- tronless mobile elements, Nature601, 440 (2022)
2022
-
[54]
P. J. Lehner, Silencing by the hush epigenetic transcrip- tional repressor complex, Annual Review of Biochemistry 94, 361 (2025)
2025
-
[55]
R. C. Allshire and H. D. Madhani, Ten principles of heterochromatin formation and function, Nature reviews Molecular cell biology19, 229 (2018)
2018
-
[56]
M. D. Aljohani, S. El Mouridi, M. Priyadarshini, A. M. Vargas-Velazquez, and C. Frøkjær-Jensen, Engineering rules that minimize germline silencing of transgenes in simple extrachromosomal arrays in c. elegans, Nature Communications11, 6300 (2020)
2020
-
[57]
Van Meter, M
M. Van Meter, M. Kashyap, S. Rezazadeh, A. J. Geneva, T. D. Morello, A. Seluanov, and V. Gorbunova, Sirt6 re- presses line1 retrotransposons by ribosylating kap1 but this repression fails with stress and age, Nature commu- nications5, 5011 (2014)
2014
-
[58]
Korotkov, A
A. Korotkov, A. Seluanov, and V. Gorbunova, Sirtuin 6: linking longevity with genome and epigenome stability, Trends in cell biology31, 994 (2021)
2021
-
[59]
Gorbunova, A
V. Gorbunova, A. Seluanov, P. Mita, W. McKerrow, D. Feny¨ o, J. D. Boeke, S. B. Linker, F. H. Gage, J. A. Kreiling, A. P. Petrashen,et al., The role of retrotrans- posable elements in ageing and age-associated diseases, Nature596, 43 (2021)
2021
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.