Effects of spatial environmental noise on evolution of cooperation
Pith reviewed 2026-06-26 18:53 UTC · model grok-4.3
The pith
Annealed environmental noise shifts phase boundaries upward to expand both the cooperative regime and extinction region in a spatial game, while quenched noise leaves boundaries nearly unchanged.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
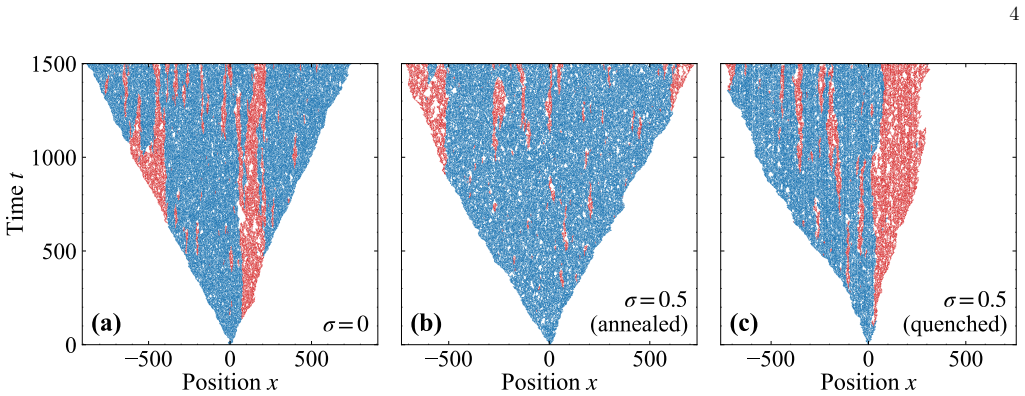
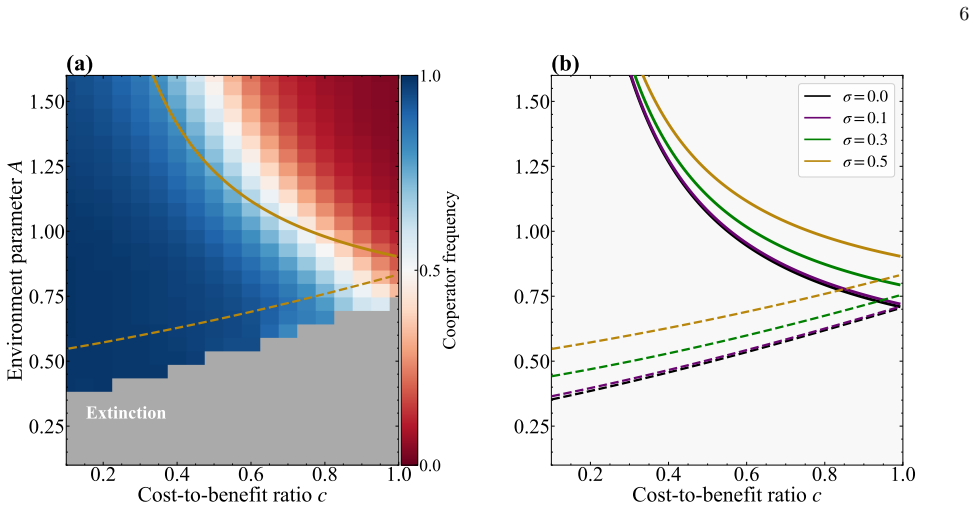
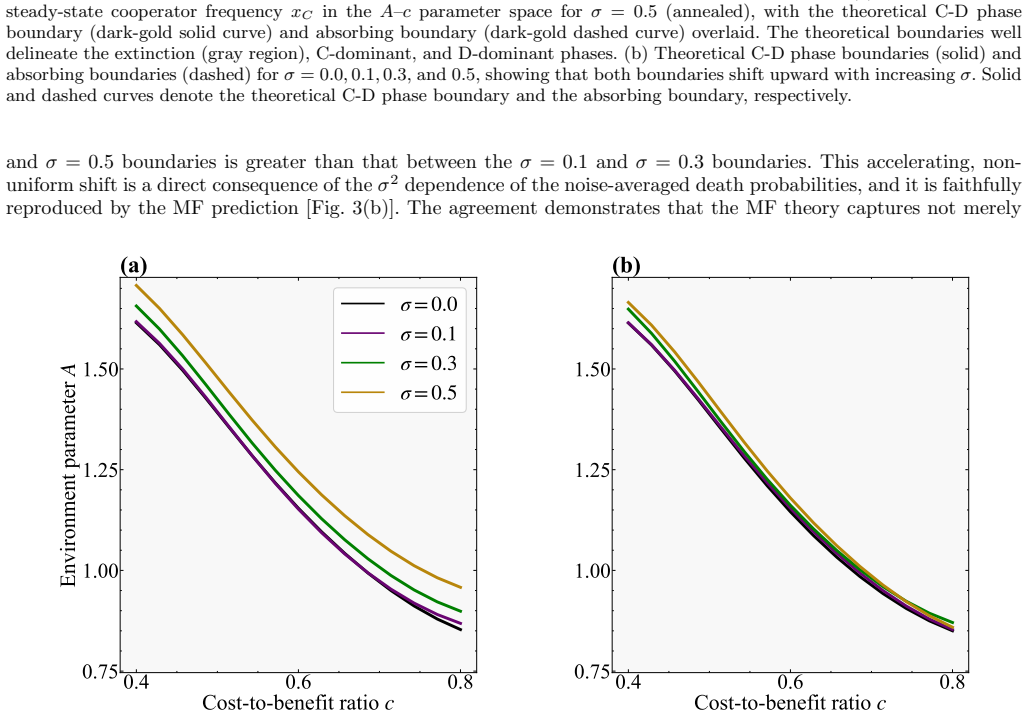
In a spatial evolutionary game on a one-dimensional lattice with vacancies, annealed noise in environmental quality increases the intensity of fluctuations that shift the cooperator-defector phase boundary and the absorbing boundary upward, thereby expanding both the regime where cooperation persists and the region of extinction, as confirmed by mean-field theory and simulations; quenched noise, by contrast, leaves the phase boundary nearly unchanged and has only a weak effect on cooperator frequency.
What carries the argument
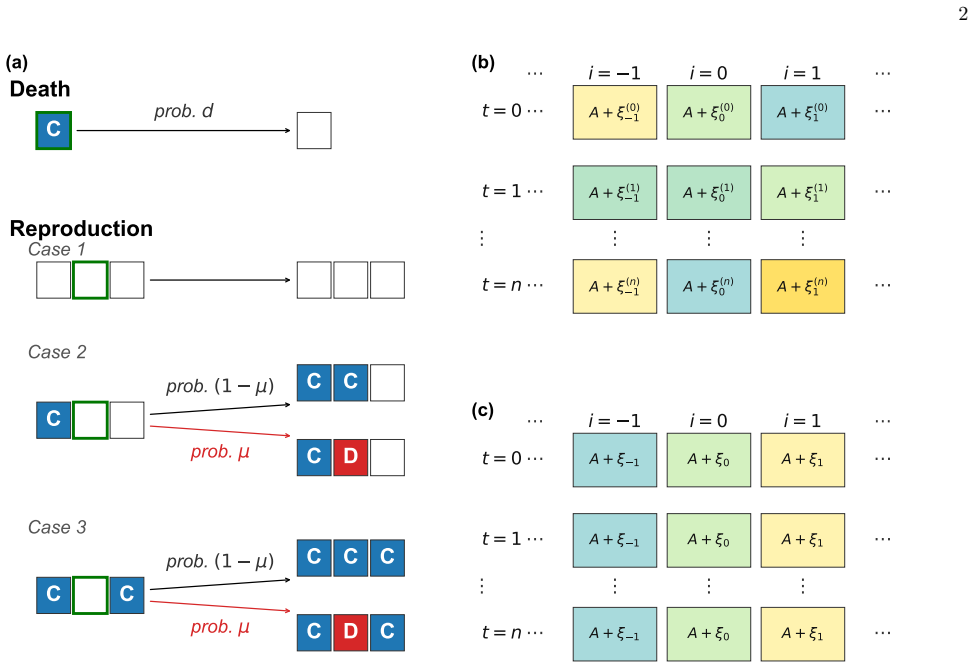
Mean-field averaging of noise-dependent death probabilities for annealed noise, contrasted with fixed per-site values for quenched noise, applied within a one-dimensional lattice model with vacancies that promotes cooperation through spatial selection.
If this is right
- Increasing annealed noise intensity expands the parameter space supporting cooperation.
- The same increase expands the region of extinction.
- Quenched noise leaves the cooperator-defector phase boundary nearly unchanged at all noise levels.
- Temporal fluctuations rather than static spatial heterogeneity drive the observed shifts in cooperative phase structure.
Where Pith is reading between the lines
- Populations subject to daily or seasonal environmental changes could maintain cooperation over wider parameter ranges than those in fixed heterogeneous habitats.
- Testing the same annealed versus quenched distinction on two-dimensional lattices would check whether the boundary shifts depend on the dimensionality of spatial structure.
- High annealed noise might raise overall extinction risk, suggesting a trade-off between cooperation and population persistence under fluctuating conditions.
- The averaging technique for death probabilities could be adapted to other models of fluctuating selection in evolutionary dynamics.
Load-bearing premise
The mean-field replacement of noise-dependent death probabilities with distribution averages accurately captures the annealed stochastic dynamics, and the one-dimensional lattice with vacancies captures the essential spatial selection without higher-dimensional effects altering the noise impact.
What would settle it
Direct stochastic simulations on the lattice that show no upward shift in the cooperator-defector or absorbing boundaries as annealed noise intensity increases would falsify the mean-field prediction.
Figures
read the original abstract
We investigate the effects of environmental noise on cooperation in a spatial evolutionary game model with variable population size. Building on a one-dimensional lattice model in which vacancies promote cooperation through spatial selection, we add random noise to the environmental quality parameter and consider two distinct types: annealed noise, where the environmental quality fluctu ates independently at each site and each time step, and quenched noise, where each site is assigned a permanently fixed random value. For annealed noise, we develop a mean-field theory by replacing the noise-dependent death probabilities with their distribution averages, and find that increasing the noise intensity shifts both the cooperator-defector phase boundary and the absorbing boundary upward in the parameter space, simultaneously expanding the cooperative regime and the extinc tion region. These predictions are confirmed by numerical simulations. In contrast, quenched noise leaves the phase boundary nearly unchanged across all noise levels, exerting only a weak effect on cooperator frequency. Together, these results demonstrate that temporal fluctuations, rather than static spatial heterogeneity, are the primary driver of noise-induced shifts in the cooperative phase structure.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript examines how annealed (temporally fluctuating) versus quenched (spatially fixed) environmental noise affects the phase structure of cooperation in a one-dimensional lattice evolutionary game with vacancies and variable population size. For annealed noise, a mean-field closure replaces each site's noise-dependent death probability with its ensemble average, predicting that higher noise intensity shifts both the cooperator-defector boundary and the absorbing boundary upward, thereby enlarging the cooperative region while also expanding the extinction region; these shifts are reported to be confirmed by direct lattice simulations. Quenched noise is found to leave the phase boundary essentially unchanged and to exert only weak effects on cooperator frequency. The central contrast is that temporal fluctuations, rather than static heterogeneity, drive the reported changes in cooperative phase structure.
Significance. If the mean-field predictions and their simulation confirmation hold, the work provides a clear demonstration that the temporal character of environmental noise can simultaneously promote cooperation and increase extinction risk in spatially structured populations, while static spatial heterogeneity does not. The explicit comparison of annealed and quenched cases, together with the availability of both an analytic closure and numerical checks, supplies a useful benchmark for future studies of fluctuating environments in evolutionary games.
major comments (2)
- [Mean-field theory] Mean-field theory section (description of averaging death probabilities): the closure replaces p_d(ε) with ⟨p_d⟩ at every site, discarding possible couplings between local density fluctuations and independent site-time noise realizations on the 1D lattice. Because reproduction and death remain strictly nearest-neighbor, this approximation is load-bearing for the claimed boundary shifts; the manuscript does not report a quantitative measure (e.g., relative error in boundary location or cooperator density) of the closure discrepancy across the scanned noise intensities.
- [Numerical simulations] Numerical simulations section: although the abstract states that simulations confirm the mean-field shifts, no table or figure directly overlays the analytic boundary locations against the simulated ones as a function of noise intensity, nor is an explicit check provided that the discrepancy remains small near the phase boundaries where spatial correlations are expected to be strongest.
minor comments (2)
- [Abstract] Abstract contains typographical errors: “fluctu ates” and “extinc tion”.
- [Model definition] Notation for the environmental quality parameter and its distribution should be introduced once in the model section and used consistently thereafter.
Simulated Author's Rebuttal
We thank the referee for the careful reading and for highlighting the value of the annealed-versus-quenched comparison. We address the two major comments below and will incorporate the requested quantitative checks in a revised manuscript.
read point-by-point responses
-
Referee: [Mean-field theory] Mean-field theory section (description of averaging death probabilities): the closure replaces p_d(ε) with ⟨p_d⟩ at every site, discarding possible couplings between local density fluctuations and independent site-time noise realizations on the 1D lattice. Because reproduction and death remain strictly nearest-neighbor, this approximation is load-bearing for the claimed boundary shifts; the manuscript does not report a quantitative measure (e.g., relative error in boundary location or cooperator density) of the closure discrepancy across the scanned noise intensities.
Authors: We agree that the mean-field closure is an approximation whose accuracy should be quantified. While the analytic boundaries are derived under the stated averaging and the simulations reproduce the predicted upward shifts, we did not supply explicit error metrics. In the revision we will add a supplementary figure (or table) that reports the relative discrepancy between mean-field and simulated cooperator densities and boundary locations for each noise intensity examined. This will allow readers to assess the approximation’s performance directly. revision: yes
-
Referee: [Numerical simulations] Numerical simulations section: although the abstract states that simulations confirm the mean-field shifts, no table or figure directly overlays the analytic boundary locations against the simulated ones as a function of noise intensity, nor is an explicit check provided that the discrepancy remains small near the phase boundaries where spatial correlations are expected to be strongest.
Authors: We accept the observation. Although the text states that simulations confirm the shifts, we did not present overlaid boundary curves or a dedicated near-boundary error analysis. The revision will include a new figure that superimposes the mean-field phase boundaries on the simulation results across the scanned noise intensities, together with a short discussion of the discrepancy in the vicinity of the critical lines. revision: yes
Circularity Check
No significant circularity; mean-field averaging and lattice simulations are independent
full rationale
The derivation begins from an explicit spatial lattice model with local reproduction/death rules and vacancies. The annealed-noise mean-field step replaces each site's p_d(ε) with its ensemble average ⟨p_d⟩ inside the existing rate equations; this is an approximation, not a redefinition of the target quantities. The resulting phase-boundary shifts are then checked by direct Monte-Carlo simulation of the original stochastic process, which is an external falsification test rather than a tautology. No fitted parameters are relabeled as predictions, no self-citation supplies a uniqueness theorem, and no ansatz is smuggled in. The reported expansion of the cooperative regime therefore rests on content that is not equivalent to its inputs by construction.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Death probabilities depend on environmental quality in a manner that permits replacement by distribution averages for annealed noise.
Reference graph
Works this paper leans on
-
[1]
Sigmund,The Calculus of Selfishness(Princeton University Press, Princeton, 2010)
K. Sigmund,The Calculus of Selfishness(Princeton University Press, Princeton, 2010)
2010
-
[2]
M. A. Nowak, Science314, 1560–1563 (2006)
2006
-
[3]
Axelrod and W
R. Axelrod and W. D. Hamilton, Science211, 1390–1396 (1981)
1981
-
[4]
J. M. Smith,Evolution and the Theory of Games(Cambridge University Press, Cambridge, 1982)
1982
-
[5]
M. A. Nowak,Evolutionary Dynamics(Harvard University Press, Cambridge, 2006)
2006
-
[6]
M. A. Nowak and R. M. May, Nature359, 826–829 (1992)
1992
-
[7]
Szab´ o and G
G. Szab´ o and G. F´ ath, Phys. Rep.446, 97–216 (2007)
2007
-
[8]
M. A. Nowak and K. Sigmund, Science303, 793–799 (2004)
2004
-
[9]
J. A. Fletcher and M. Doebeli, Proc. R. Soc. B276, 13–19 (2009)
2009
-
[10]
Ohtsuki, C
H. Ohtsuki, C. Hauert, E. Lieberman, and M. A. Nowak, Nature441, 502–505 (2006)
2006
-
[11]
F. C. Santos, J. M. Pacheco, and T. Lenaerts, Proc. Natl. Acad. Sci. USA103, 3490–3494 (2006). 8
2006
-
[12]
Szab´ o and C
G. Szab´ o and C. T˝ oke, Phys. Rev. E58, 69–73 (1998)
1998
-
[13]
Perc and A
M. Perc and A. Szolnoki, Biosystems99, 109–125 (2010)
2010
-
[14]
M. Perc, J. J. G´ omez-Garde˜ nes, A. Szolnoki, L. M. Flor´ ıa, and Y. Moreno, J. R. Soc. Interface10, 20120997 (2013)
2013
-
[15]
D´ ebarre, C
F. D´ ebarre, C. Hauert, and M. Doebeli, Nat. Commun.5, 3409 (2014)
2014
-
[16]
Melbinger, J
A. Melbinger, J. Cremer, and E. Frey, Phys. Rev. Lett.105, 178101 (2010)
2010
-
[17]
Hauert, M
C. Hauert, M. Holmes, and M. Doebeli, Proc. R. Soc. B273, 2565–2571 (2006)
2006
-
[18]
G. W. A. Constable, T. Rogers, A. J. McKane, and C. E. Tarnita, Proc. Natl. Acad. Sci. USA113, E4745–E4754 (2016)
2016
-
[19]
Huang, C
W. Huang, C. Hauert, and A. Traulsen, Proc. Natl. Acad. Sci. USA112, 9064–9069 (2015)
2015
-
[20]
Szab´ o, J
G. Szab´ o, J. Vukov, and A. Szolnoki, Phys. Rev. E72, 047107 (2005)
2005
-
[21]
Hinrichsen, Adv
H. Hinrichsen, Adv. Phys.49, 815–958 (2000)
2000
-
[22]
S. Chae, N. Lee, S. K. Baek, and H.-C. Jeong, Phys. Rev. E103, 032114 (2021)
2021
-
[23]
H. J. Park, C. Hilbe, M. A. Nowak, B. J. Kim, and H.-C. Jeong, J. Theor. Biol.575, 111629 (2023)
2023
-
[24]
Chesson, Annu
P. Chesson, Annu. Rev. Ecol. Syst.31, 343–366 (2000)
2000
-
[25]
Hilbe, ˇS
C. Hilbe, ˇS. ˇSimsa, K. Chatterjee, and M. A. Nowak, Nature559, 246–249 (2018)
2018
-
[26]
Ashcroft, P
P. Ashcroft, P. M. Altrock, and T. Galla, J. R. Soc. Interface11, 20140663 (2014)
2014
-
[27]
J. W. Baron and T. Galla, R. Soc. Open Sci.5, 172176 (2018)
2018
-
[28]
C. P. Roca, J. A. Cuesta, and A. S´ anchez, Phys. Life Rev.6, 208–249 (2009)
2009
-
[29]
A. R. Tilman, J. B. Plotkin, and E. Akcay, Nat. Commun.11, 915 (2020)
2020
-
[30]
M. H. Vainstein and J. J. Arenzon, Phys. Rev. E64, 051905 (2001)
2001
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.