When do correlations reflect biological similarity in ecological dynamics?
Pith reviewed 2026-06-26 06:18 UTC · model grok-4.3
The pith
Abundance correlations do not reflect niche overlap in stochastic Lotka-Volterra models even when stochastic forcing matches biological similarity.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
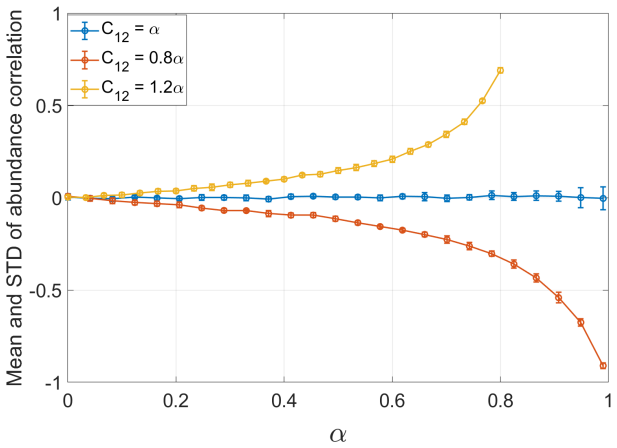
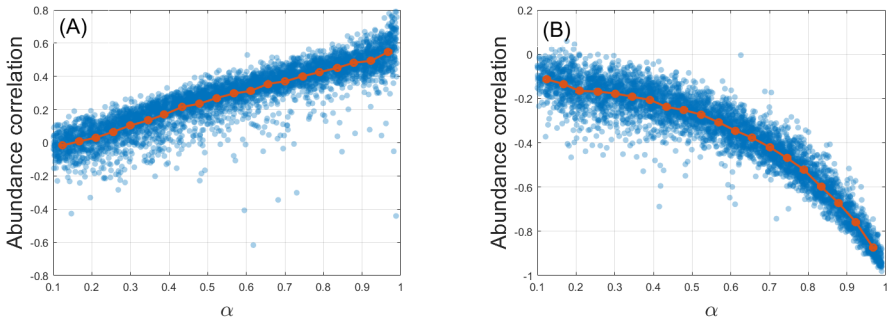
Requiring that stochastic forcing on different species be correlated in proportion to their biological similarity cannot, in general, be satisfied within the stochastic Lotka-Volterra framework; even in the special cases where it can be satisfied, the resulting abundance correlations carry no information about niche overlap. Consumer-resource models supply a natural setting in which stochasticity can be biologically grounded, yet the interpretation of abundance correlations in those models depends on the pathway through which noise enters the system.
What carries the argument
The mapping from biologically proportional stochastic forcing to abundance correlations, which fails to encode niche overlap inside the stochastic Lotka-Volterra framework.
If this is right
- Abundance correlations cannot be interpreted as measures of niche overlap inside stochastic Lotka-Volterra models.
- Consumer-resource models are required if stochasticity is to be introduced in a manner proportional to biological similarity.
- Even inside consumer-resource models the quantity encoded by abundance correlations changes with the route taken by the noise.
- Studies that treat microbial abundance correlations as direct read-outs of interaction strength rest on modeling assumptions that are not generally valid.
Where Pith is reading between the lines
- Empirical time-series studies that rely on abundance correlations to infer community structure may need to specify the underlying model before interpreting their results.
- Alternative model classes, such as explicit resource dynamics with separate noise sources, could be tested against the same microbial datasets to see whether they recover known niche relationships.
- The paper leaves open whether a hybrid modeling approach could restore a direct link between forcing correlations and observed abundance correlations.
Load-bearing premise
That stochastic forcing acting on different species must be correlated in proportion to their biological similarity in order to be biologically grounded.
What would settle it
A numerical simulation of a stochastic Lotka-Volterra system in which the noise covariance matrix is set exactly proportional to niche overlap and the resulting abundance correlation matrix is checked for any systematic relation to the overlap matrix.
Figures
read the original abstract
The structure of competitive ecological communities is shaped by the strength of interactions between species, which in turn reflects their biological similarity. At the same time, the stochastic forcing that drives abundance fluctuations is itself biologically grounded: species that are more similar may be expected to respond more similarly to environmental variation. This motivates the increasingly common use of correlations in abundance time series, particularly in microbial communities, as proxies for biological similarity or niche overlap. Here we analyze the relation between biological similarity and abundance correlations in stochastic community models. We require that the stochastic forcing acting on different species be correlated in proportion to their biological similarity, and ask how such forcing is reflected in abundance correlations. We show that this requirement cannot, in general, be satisfied within the widely used stochastic Lotka-Volterra framework, and that even when it is, abundance correlations carry no information about niche overlap. In contrast, consumer-resource models provide a natural framework for biologically grounded stochasticity. In this setting, however, the interpretation of abundance correlations depends strongly on the pathway through which noise enters the system: direct forcing of consumers and resource-mediated fluctuations encode different biological quantities. These results have implications both for the modeling of stochastic ecological communities and for understanding what can, and cannot, be inferred from correlations in community time series.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper analyzes whether abundance correlations in stochastic ecological models can serve as proxies for biological similarity or niche overlap. It imposes the requirement that stochastic forcing correlations must be proportional to biological similarity and demonstrates that this cannot generally be realized within the stochastic Lotka-Volterra (SLV) framework; even when realizable, the resulting abundance correlations are independent of overlap. Consumer-resource (CR) models are shown to accommodate biologically grounded stochasticity, but the mapping from noise to abundance correlations depends on whether noise acts directly on consumers or is mediated by resources.
Significance. If the analytical results hold, the work cautions against routine use of time-series correlations as measures of niche overlap, especially in microbial ecology. The explicit contrast between SLV and CR frameworks, together with the pathway dependence in the latter, supplies a concrete basis for model choice and for interpreting what can versus cannot be inferred from community time series. The purely analytical character of the argument is a strength.
minor comments (2)
- [§2] §2: the precise definition of the noise-correlation matrix (proportional to overlap) should be stated as an explicit equation before the main theorems, to make the 'requirement' unambiguous.
- The transition from the general SDE to the specific SLV and CR forms would benefit from a short table listing the drift and diffusion terms side-by-side.
Circularity Check
No significant circularity identified
full rationale
The paper derives its central claim by imposing the modeling requirement that stochastic forcing correlations equal biological similarity (niche overlap) and then solving the resulting stochastic Lotka-Volterra SDEs to show that this requirement is generally unrealizable and that abundance correlations are independent of overlap when it is realizable. This step is a direct algebraic consequence of the SDE structure under the stated premise and does not reduce to any fitted parameter, self-defined quantity, or self-citation chain. The contrast with consumer-resource models likewise follows from explicit comparison of noise-entry pathways without invoking prior author results as load-bearing uniqueness theorems. No quoted equation or derivation step matches any of the six enumerated circularity patterns; the analysis remains self-contained against the model equations themselves.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Stochastic forcing acting on different species is correlated in proportion to their biological similarity
Reference graph
Works this paper leans on
-
[1]
The coefficientα ij reflects the extent to which species depend on similar resources. Accordingly, the correlations between their environmental responses are expected to increase withα ij: species with larger niche overlap should experience more similar fluctuations in their growth rates
-
[2]
It is therefore natural to expect that external environmental factors, such as temperature or pH, will affect species in a similar manner whenαij is large
Even setting aside resource dynamics, the interaction coefficientαij often reflects bio- logical similarity between species, for example phylogenetic relatedness. It is therefore natural to expect that external environmental factors, such as temperature or pH, will affect species in a similar manner whenαij is large. Wearethereforeledtomodelstochasticityi...
-
[3]
Russell Lande, Steinar Engen, and Bernt-Erik Saether,Stochastic population dynamics in ecology and conservation(Oxford University Press, 2003)
2003
-
[4]
Effects of temporal environmental stochasticity on species richness: a mechanistic unification spanning weak to strong temporal correlations,
Tak Fung, James P O’Dwyer, and Ryan A Chisholm, “Effects of temporal environmental stochasticity on species richness: a mechanistic unification spanning weak to strong temporal correlations,” Oikos2022, e08667 (2022)
2022
-
[5]
Ecological communities with lotka-volterra dynamics,
Guy Bunin, “Ecological communities with lotka-volterra dynamics,” Physical Review E95, 042414 (2017)
2017
-
[6]
Generic assembly patterns in complex ecological communities,
Matthieu Barbier, Jean-François Arnoldi, Guy Bunin, and Michel Loreau, “Generic assembly patterns in complex ecological communities,” Proceedings of the National Academy of Sciences 115, 2156–2161 (2018)
2018
-
[7]
A neutral theory with envi- ronmental stochasticity explains static and dynamic properties of ecological communities,
Michael Kalyuzhny, Ronen Kadmon, and Nadav M Shnerb, “A neutral theory with envi- ronmental stochasticity explains static and dynamic properties of ecological communities,” Ecology letters18, 572–580 (2015)
2015
-
[8]
Theory of time-averaged neutral dynamics with envi- ronmental stochasticity,
Matan Danino and Nadav M Shnerb, “Theory of time-averaged neutral dynamics with envi- ronmental stochasticity,” Physical Review E97, 042406 (2018)
2018
-
[9]
Chaotic turnover of rare and abundant species in a strongly interacting model community,
Emil Mallmin, Arne Traulsen, and Silvia De Monte, “Chaotic turnover of rare and abundant species in a strongly interacting model community,” Proceedings of the National Academy of Sciences121, e2312822121 (2024)
2024
-
[10]
Many-species ecological fluctuations as a jump process from the brink of extinction,
Thibaut Arnoulx de Pirey and Guy Bunin, “Many-species ecological fluctuations as a jump process from the brink of extinction,” Physical Review X14, 011037 (2024)
2024
-
[11]
Macroecologicallawsdescribevariationanddiversityinmicrobialcommunities,
JacopoGrilli,“Macroecologicallawsdescribevariationanddiversityinmicrobialcommunities,” Nature communications11, 1–11 (2020)
2020
-
[12]
In- teractions between strains govern the eco-evolutionary dynamics of microbial communities,
Akshit Goyal, Leonora S Bittleston, Gabriel E Leventhal, Lu Lu, and Otto X Cordero, “In- teractions between strains govern the eco-evolutionary dynamics of microbial communities,” Elife11, e74987 (2022)
2022
-
[13]
Environmental fluctuations explain the universal decay of species-abundance correlations with phylogenetic distance,
Matteo Sireci, Miguel A Muñoz, and Jacopo Grilli, “Environmental fluctuations explain the universal decay of species-abundance correlations with phylogenetic distance,” Proceedings of the National Academy of Sciences120, e2217144120 (2023). 18
2023
-
[14]
Microbial functional guilds respond cohesively to rapidly fluctuating environments,
Kyle Crocker, Abigail Skwara, Rathi Kannan, Arvind Murugan, and Seppe Kuehn, “Microbial functional guilds respond cohesively to rapidly fluctuating environments,” bioRxiv (2025)
2025
-
[15]
Inferring resource competition in microbial communities from time series,
Xiaowen Chen, Kyle Crocker, Seppe Kuehn, Aleksandra M Walczak, and Thierry Mora, “Inferring resource competition in microbial communities from time series,” PRX Life3, 023019 (2025)
2025
-
[16]
Biodiversity and ecosystem stability: a synthesis of underlying mechanisms,
Michel Loreau and Claire De Mazancourt, “Biodiversity and ecosystem stability: a synthesis of underlying mechanisms,” Ecology letters16, 106–115 (2013)
2013
-
[17]
Front propagation into unstable states,
Wim Van Saarloos, “Front propagation into unstable states,” Physics reports386, 29–222 (2003)
2003
-
[18]
Species competition and predation,
Peter Chesson, “Species competition and predation,” inEcological systems(Springer, 2013) pp. 223–256
2013
-
[19]
Intuitive and broadly applicable definitions of niche and fitness differences,
Jurg W Spaak and Frederik De Laender, “Intuitive and broadly applicable definitions of niche and fitness differences,” Ecology Letters23, 1117–1128 (2020)
2020
-
[20]
Macarthur’s consumer-resource model: a rosetta stone for competitive interactions,
Jawad Sakarchi and Rachel M Germain, “Macarthur’s consumer-resource model: a rosetta stone for competitive interactions,” The American Naturalist205, 306–326 (2025)
2025
-
[21]
Niche overlap is not enough: Same overlap, opposite dynamics,
Akiva Goldberg, Oshrit Shtossel, Yoram Louzoun, and Nadav M Shnerb, “Niche overlap is not enough: Same overlap, opposite dynamics,” bioRxiv (2025), accepted, Ecology letters
2025
-
[22]
Enzymic analysis of the crabtree effect in glucose-limited chemostat cultures of saccharomyces cerevisiae,
E. Postma, C. Verduyn, W. A. Scheffers, and J. P. Van Dijken, “Enzymic analysis of the crabtree effect in glucose-limited chemostat cultures of saccharomyces cerevisiae,” Applied and Environmental Microbiology55, 468–477 (1989)
1989
-
[23]
Temperature dependence of energy conversion efficiency in insect larvae,
J. D. Bjørge, J. Overgaard, and H. Malte, “Temperature dependence of energy conversion efficiency in insect larvae,” Journal of Thermal Biology74, 252–258 (2018)
2018
-
[24]
Overflow metabolism in escherichia coli results from efficient proteome allocation,
M. Basan, S. Hui, H. Okano, Z. Zhang, Y. Shen, J. R. Williamson, and T. Hwa, “Overflow metabolism in escherichia coli results from efficient proteome allocation,” Nature528, 99–104 (2015)
2015
-
[25]
Body size, energy metabolism and lifespan,
J. R. Speakman, “Body size, energy metabolism and lifespan,” Journal of Experimental Biology 208, 1717–1730 (2005)
2005
-
[26]
The effect of prey and predator densities on wolf predation,
J. A. Vucetich, R. O. Peterson, and C. L. Schaefer, “The effect of prey and predator densities on wolf predation,” Ecology83, 3003–3013 (2004)
2004
-
[27]
Owen-Smith,Megaherbivores: The Influence of Very Large Body Size on Ecology(Cam- bridge University Press, 1988)
N. Owen-Smith,Megaherbivores: The Influence of Very Large Body Size on Ecology(Cam- bridge University Press, 1988). 19
1988
-
[28]
Scatter hoarding by rodents and seed predation: a spatially explicit model of survival,
P. A. Jansen, M. Bartholomeus, and F. Bongers, “Scatter hoarding by rodents and seed predation: a spatially explicit model of survival,” Ecological Modelling175, 417–430 (2004)
2004
-
[29]
Scent-marking by male mammals: cheat-proof signals to competitors and mates,
L. M. Gosling and S. C. Roberts, “Scent-marking by male mammals: cheat-proof signals to competitors and mates,” Advances in the Study of Behavior30, 169–217 (2001)
2001
-
[30]
Ecosystem stability relies on diversity difference between trophic levels,
Yizhou Liu, Jiliang Hu, and Jeff Gore, “Ecosystem stability relies on diversity difference between trophic levels,” Proceedings of the National Academy of Sciences121, e2416740121 (2024)
2024
-
[31]
Stability criteria for the consumption and exchange of essential resources,
Theo Gibbs, Yifan Zhang, Zachary R Miller, and James P O’Dwyer, “Stability criteria for the consumption and exchange of essential resources,” PLoS computational biology18, e1010521 (2022)
2022
-
[32]
Phase transition to chaos in com- plex ecosystems with nonreciprocal species-resource interactions,
Emmy Blumenthal, Jason W Rocks, and Pankaj Mehta, “Phase transition to chaos in com- plex ecosystems with nonreciprocal species-resource interactions,” Physical review letters132, 127401 (2024)
2024
-
[33]
Species competition and predation,
Peter Chesson, “Species competition and predation,” inEcological systems: Selected entries from the encyclopedia of sustainability science and technology(Springer, 2012) pp. 223–256
2012
-
[34]
A tiny fraction of all species forms most of nature: Rarity as a sticky state,
Egbert H van Nes, Diego GF Pujoni, Sudarshan A Shetty, Gerben Straatsma, Willem M de Vos, and Marten Scheffer, “A tiny fraction of all species forms most of nature: Rarity as a sticky state,” Proceedings of the National Academy of Sciences121, e2221791120 (2024)
2024
-
[35]
Gene H Golub and Charles F Van Loan,Matrix computations(JHU press, 2013)
2013
-
[36]
Will a large complex system be stable?
Robert M May, “Will a large complex system be stable?” Nature238, 413–414 (1972)
1972
-
[37]
Communitiesas cliques,
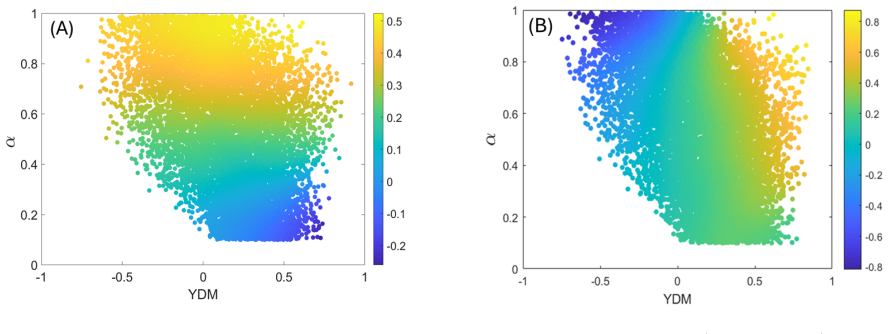
Yael Fried, DavidA Kessler, and Nadav MShnerb, “Communitiesas cliques,” Scientific reports 6, 35648 (2016). 20 Appendix A: The yield-depletion mismatch (YDM) In the main text, we emphasized that when stochasticity is transmitted through resource fluctuations, abundance correlations between consumers are not primarily governed by niche overlap alone. Inste...
2016
-
[38]
At the level of the reduced Lotka-Volterra description, their level of competition is encoded by the effective interaction coefficientαij
Resource-mediated correlations are controlled by yield-depletion mismatch Consider two consumer speciesiandjrelying on a common pool of resources. At the level of the reduced Lotka-Volterra description, their level of competition is encoded by the effective interaction coefficientαij. In a consumer-resource systemαij captures their niche overlap between t...
-
[39]
(1) of the main 21 text): ∆γ = 1−cosθ i,j(γ)≡1− ⃗ γi ·⃗ γj |⃗ γi||⃗ γj| ∆λ = 1−cosθ i,j(λ)≡1− ⃗λi · ⃗λj |⃗λi||⃗λj|
Definition We distinguish between two aspects of functional dissimilarity, namely yield dissimilarity ∆γ and depletion dissimilarity∆λ, defined through the cosine distance between the relevant rows of the yield matrixΓ, and columns of the depletion matrixΛ(see Eq. (1) of the main 21 text): ∆γ = 1−cosθ i,j(γ)≡1− ⃗ γi ·⃗ γj |⃗ γi||⃗ γj| ∆λ = 1−cosθ i,j(λ)≡1...
-
[40]
When∆ γ >∆ λ, the two species differ more in yield than in depletion, soD <0
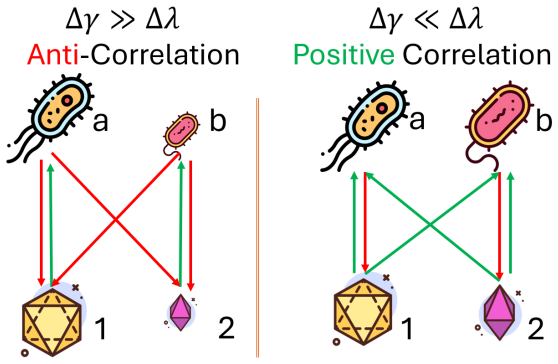
Interpretation The sign ofDhas a simple qualitative meaning. When∆ γ >∆ λ, the two species differ more in yield than in depletion, soD <0. In that case, a fluctuation in resource availability may benefit one species much more than the other, while both species still impose similar pressure on the resource pool. This tends to generate negative abundance co...
-
[41]
Relation to previous work The role of YDM in the present paper is conceptually distinct from the role it played in [19]. There, the main question was whether abundance correlations can be predicted from niche overlap in a stochastic consumer-resource system, and the answer was negative: the relevant predictor was instead the yield-depletion mismatch. Here...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.