Far-from-equilibrium travelling pulses in sloped semi-arid environments driven by autotoxicity effects
Pith reviewed 2026-05-24 01:25 UTC · model grok-4.3
The pith
An extension of the Klausmeier model shows autotoxicity can produce travelling vegetation pulses on slopes.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
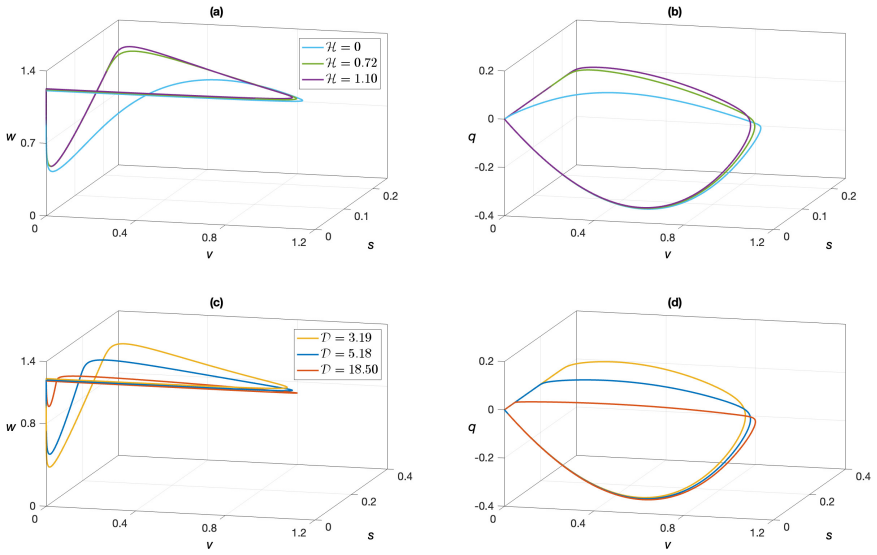
In the extended Klausmeier model that incorporates autotoxicity, travelling pulse solutions exist and can be proven by constructing corresponding homoclinic orbits in the associated four-dimensional ordinary differential equation system. This construction is carried out via geometric singular perturbation theory once a scaling analysis places the system in an appropriate singularly perturbed regime. The resulting solutions match numerical simulations and allow extraction of biological observations on the role of autotoxicity in vegetation dynamics.
What carries the argument
Construction of homoclinic orbits in the four-dimensional travelling-wave system obtained after scaling the model equations, using geometric singular perturbation theory to handle the fast-slow decomposition.
If this is right
- Travelling pulses persist under autotoxicity in sloped semi-arid settings.
- Pulse speed and width are controlled by the strength of the toxicity feedback.
- Autotoxicity can shift stationary patterns into moving ones.
- The analytic pulses agree quantitatively with direct numerical simulations of the PDE.
- Biological conclusions follow about how plant-produced toxins affect vegetation movement on slopes.
Where Pith is reading between the lines
- The same scaling and homoclinic construction technique could be tested on other extensions of the Klausmeier model that include different feedback mechanisms.
- Two-dimensional versions of the model might admit analogous travelling stripes whose stability could be checked numerically.
- The role of autotoxicity in preventing or enabling pattern invasion into bare soil could be explored by varying the toxicity parameter across the existence threshold.
Load-bearing premise
The model parameters admit a scaling that places the travelling-wave system in a singularly perturbed regime where the fast-slow decomposition remains valid.
What would settle it
Numerical integration of the scaled four-dimensional travelling-wave system that fails to locate the predicted homoclinic orbits, or constructed orbits whose profiles deviate substantially from simulated pulse solutions.
Figures









read the original abstract
In this work, an extension of the 1D Klausmeier model that accounts for the toxicity compounds is considered and the occurrence of travelling stripes is investigated. Numerical simulations are firstly conducted to capture the qualitative behaviours of the pulse-type solutions and, then, geometric singular perturbation theory is used to prove the existence of such travelling pulses by constructing the corresponding homoclinic orbits in the associated 4-dimensional system. A scaling analysis on the investigated model is performed to identify the asymptotic scaling regime in which travelling pulses can be constructed. Biological observations are extracted from the analytical results and the role of autotoxicity in travelling patterns is emphasized. Finally, the analytically constructed solutions are compared with the numerical ones, leading to a good agreement that confirms the validity of the conducted analysis. Numerical investigations are also carried out in order to gain additional information on vegetation dynamics.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript extends the 1D Klausmeier vegetation model by adding an autotoxicity term and studies travelling pulse (stripe) solutions on slopes. Numerical simulations first illustrate the pulses; a scaling analysis then reduces the PDE to a 4D fast-slow ODE system in a specific asymptotic regime, after which geometric singular perturbation theory is invoked to prove existence by constructing a homoclinic orbit. Biological conclusions are drawn from the analytic pulses, and the constructed solutions are compared with the numerics, showing good agreement.
Significance. If the GSPT construction is valid, the work supplies the first rigorous existence proof for toxicity-driven travelling pulses in an extended Klausmeier system, together with explicit parameter regimes and direct numerical validation. This strengthens the mathematical foundation for pattern-formation models in semi-arid ecology and isolates the mechanistic role of autotoxicity.
major comments (1)
- [Scaling analysis and GSPT construction] Scaling analysis and GSPT construction: the manuscript identifies an asymptotic regime that yields a 4D system, but does not explicitly verify normal hyperbolicity of the critical manifold (eigenvalues of the fast Jacobian) or the transversality conditions required for the slow-manifold matching that assembles the homoclinic orbit. These verifications are load-bearing for the existence claim; without them the application of Fenichel theory and the subsequent geometric construction remain incomplete.
Simulated Author's Rebuttal
We thank the referee for their careful reading of the manuscript and for highlighting the need for explicit verification of key technical conditions in the GSPT construction. We address the major comment below and will incorporate the necessary additions in a revised version.
read point-by-point responses
-
Referee: Scaling analysis and GSPT construction: the manuscript identifies an asymptotic regime that yields a 4D system, but does not explicitly verify normal hyperbolicity of the critical manifold (eigenvalues of the fast Jacobian) or the transversality conditions required for the slow-manifold matching that assembles the homoclinic orbit. These verifications are load-bearing for the existence claim; without them the application of Fenichel theory and the subsequent geometric construction remain incomplete.
Authors: We agree that explicit verification of normal hyperbolicity and the relevant transversality conditions is required to make the application of Fenichel theory and the homoclinic construction fully rigorous. In the revised manuscript we will add a dedicated subsection that computes the eigenvalues of the fast Jacobian along the critical manifold and confirms that all eigenvalues have nonzero real parts (hence normal hyperbolicity). We will also state and verify the transversality conditions that guarantee the transverse intersection of the stable and unstable manifolds of the slow manifolds used in the matching argument. These additions will be placed immediately after the scaling analysis and before the geometric construction of the homoclinic orbit. revision: yes
Circularity Check
No circularity: GSPT existence proof is independent of inputs
full rationale
The paper extends the Klausmeier model with toxicity, performs scaling analysis to select an asymptotic regime yielding a 4D fast-slow system, and invokes geometric singular perturbation theory (external theorems on normally hyperbolic manifolds and homoclinic construction) to prove existence of travelling pulses. This chain does not reduce any claimed result to a fitted parameter, self-defined quantity, or load-bearing self-citation; the minor reference to the base Klausmeier model supplies only the starting PDE and carries no part of the existence argument. The derivation remains self-contained against external mathematical benchmarks.
Axiom & Free-Parameter Ledger
free parameters (2)
- toxicity strength parameter
- scaled slope and diffusion ratios
axioms (2)
- standard math The extended reaction-diffusion system admits a normally hyperbolic slow manifold on which the reduced flow can be analyzed.
- domain assumption The toxicity variable diffuses or decays at a rate that permits separation of time scales in the chosen asymptotic regime.
Reference graph
Works this paper leans on
-
[1]
United Nations Convention to Combat Desertfication. The Global Land Outlook . UNCCD, Bonn, second edition, 2022. ISBN 978-92-95118-53-9. URL https://www. unccd.int/resources/global-land-outlook/global-land-outlook-2nd-edition . 21
work page 2022
-
[2]
A.G. de Soyza, W.G. Whitford, J.E. Herrick, J.W. Van Zee, and K.M. Havstad. Early warning indicators of desertification: examples of tests in the Chihuahuan Desert.Journal of Arid Environments, 39(2):101–112, 1998. doi: 10.1006/jare.1998.0391
-
[3]
G. Tirabassi, J. Viebahn, V. Dakos, H.A. Dijkstra, C. Masoller, M. Rietkerk, and S.C. Dekker. Interaction network based early-warning indicators of vegetation transitions. Ecological Complexity, 19:148–157, 2014. doi: 10.1016/j.ecocom.2014.06.004
-
[4]
K. Gowda, S. Iams, and M. Silber. Signatures of human impact on self-organized veg- etation in the Horn of Africa. Scientific Reports, 8(1):3622, 2018. doi: 10.1038/s41598- 018-22075-5
-
[5]
R.M. May. Thresholds and breakpoints in ecosystems with a multiplicity of stable states. Nature, 269(5628):471–477, 1977. doi: 10.1038/269471a0
-
[6]
M. Rietkerk, F. van den Bosch, and J. van de Koppel. Site-specific properties and irreversible vegetation changes in semi-arid grazing systems. Oikos, 80:241–252, 1997. doi: 10.2307/3546592
-
[7]
P.M. Saco, M. Moreno-de las Heras, S. Keesstra, J. Baartman, O. Yetemen, and J.F. Rodr´ ıguez. Vegetation and soil degradation in drylands: nonlinear feedbacks and early warning signals. Current Opinion in Environmental Science & Health , 5:67–72, 2018. doi: 10.1016/j.coesh.2018.06.001
-
[8]
R. Bastiaansen, A. Doelman, M.B. Eppinga, and M. Rietkerk. The effect of climate change on the resilience of ecosystems with adaptive spatial pattern formation. Ecology Letters, 43:414–429, 2020. doi: 10.1111/ele.13449
-
[9]
M. Rietkerk, R. Bastiaansen, S. Banerjee, J. Van De Koppel, M. Baudena, and A. Doel- man. Evasion of tipping in complex systems through spatial pattern formation. Science, 374, 2021. doi: 10.1126/science.abj0359
-
[10]
E. Meron. Nonlinear Physics of Ecosystems. CRC Press, Boca Raton, first edition, 2015. doi: 10.1201/b18360
-
[11]
Community Ecol- ogy 8(1), 103–109 (2007)
S. Mazzoleni, G. Bonanomi, F. Giannino, M. Rietkerk, S. Dekker, and F. Zucconi. Is plant biodiversity driven by decomposition processes? an emerging new theory on plant diversity. Community Ecology, 8(1):103–109, 2007. doi: 10.1556/ComEc.8.2007.1.12
-
[12]
S. Mazzoleni, G. Bonanomi, G. Incerti, M.L. Chiusano, P. Termolino, A. Mingo, M. Sen- atore, F. Giannino, F. Carten` ı, M. Rietkerk, and V. Lanzotti. Inhibitory and toxic effects of extracellular self-dna in litter: a mechanism for negative plant–soil feedbacks? New Phytologist, 205(3):1195–1210, 2015. doi: 10.1111/nph.13121
-
[13]
Science284(5421), 1826–1828 (1999)
C.A. Klausmeier. Regular and irregular patterns in semiarid vegetation. Science, 284: 1826–1828, 1999. doi: 10.1126/science.284.5421.1826
-
[14]
R. HilleRisLambers, M. Rietkerk, F. van de Bosch, H.H.T. Prins, and H. de Kroon. Vegetation pattern formation in semi-arid grazing systems. Ecology, 82(1):50, 2001. doi: 10.1890/0012-9658(2001)082[0050:VPFISA]2.0.CO;2. 22
-
[15]
The American Naturalist 160(4), 524–530 (2002)
M. Rietkerk, M.C. Boerlijst, F. van Langevelde, R. HilleRisLambers, J. van de Koppel, H.H.T. Prins, and A. de Roos. Self-organisation of vegetation in arid ecosystems. The American Naturalist, 160(4):534, 2002. doi: 10.1086/342078
-
[16]
Journal of Mathematical Biology 51(2), 183–197 (2005)
J.A. Sherratt. An analysis of vegetation stripe formation in semi-arid landscapes. Journal of Mathematical Biology, 51:183–197, 2005. doi: 10.1007/s00285-005-0319-5
-
[17]
K. Siteur, E. Siero, M.B. Eppinga, J.D.M. Rademacher, A. Doelman, and M. Rietk- erk. Beyond turing: The response of patterned ecosystems to environmental change. Ecological Complexity, 20:81–96, 2014. doi: 10.1016/j.ecocom.2014.09.002
-
[18]
F. Carten` ı, A. Marasco, G. Bonanomi, S. Mazzoleni, M. Rietkerk, and F. Giannino. Neg- ative plant soil feedback explaining ring formation in clonal plants.Journal of Theoretical Biology, 313:153–161, 2012. doi: 10.1016/j.jtbi.2012.08.008
-
[19]
S. van der Stelt, A. Doelman, G. Hek, and J.D.M. Rademacher. Rise and fall of periodic patterns for a generalized Klausmeier-Gray-Scott model. Journal of Nonlinear Science , 23(7):39–95, 2013. doi: 10.1007/s00332-012-9139-0
-
[20]
A. Marasco, F. Giannino, and A. Iuorio. Modelling competitive interactions and plant–soil feedback in vegetation dynamics. Ricerche di Matematica, 69:553—-577, 2020. doi: 10.1007/s11587-020-00497-6
-
[21]
L. Eigentler and J.A. Sherratt. An integrodifference model for vegetation patterns in semi-arid environments with seasonality. Journal of Mathematical Biology , 81:875–904,
-
[22]
doi: 10.1007/s00285-020-01530-w
-
[23]
A. Iuorio, M.B. Eppinga, M. Baudena, F. Veerman, M. Rietkerk, and F. Giannino. How does negative plant-soil feedback across lifestages affect the spatial patterning of trees? Scientific Reports, 2023. doi: 10.1038/s41598-023-44867-0
-
[24]
A. Iuorio, M. Baudena, M.B. Eppinga, F. Giannino, M. Rietkerk, and F. Veerman. Travelling waves due to negative plant-soil feedbacks in a model including tree life-stages. Mathematical Biosciences, 2023. doi: 10.1016/j.mbs.2023.109128
-
[25]
K. Spiliotis, L. Russo, F. Giannino, and C. Siettos. Numerical bifurcation analysis of turing and symmetry broken patterns of a vegetation pde model. Preprint, 2023. doi: 10.48550/arXiv.2303.13248
-
[26]
B.J. Kealy and D.J. Wollkind. A nonlinear stability analysis of vegetative turing pattern formation for an interaction-diffusion plant-surface water model system in an arid flat en- vironment. Bulletin of Mathematical Biology , 74(4):803–833, 2012. doi: 10.1007/s11538- 011-9688-7
-
[27]
Y. Zelnik, S. Kinast, H. Yizhaq, G. Bel, and E. Meron. Regime shifts in models of dryland vegetation. Philosophical Transactions of the Royal Society A, 371(2004):20120358, 2013. doi: 10.1098/rsta.2012.0358
-
[28]
G.Q. Sun, L. Li, and Z.K. Zhang. Spatial dynamics of a vegetation model in an arid flat environment. Nonlinear Dynamics, 73(5):2207–2219, 2013. doi: 10.1007/s11071-013- 0935-3. 23
-
[29]
G. Consolo and G. Grif` o. Eckhaus instability of stationary patterns in hyperbolic reaction-diffusion models on large finite domains. Partial Differential Equations and Applications, 3:57, 2022. doi: 10.1007/s42985-022-00193-0
-
[30]
G. Grif´ o, G. Consolo, C. Curr´ o, and G. Valenti. Rhombic and hexagonal pattern for- mation in 2D hyperbolic reaction-transport systems in the context of dryland ecology. Physica D, 449:133745, 2023. doi: 10.1016/j.physd.2023.133745
-
[31]
C. Curr` o, G. Grif` o, and G. Valenti. Turing patterns in hyperbolic reaction-transport vegetation models with cross-diffusion. Chaos, Solitons & Fractals , 176:114152, 2023. doi: 10.1016/j.chaos.2023.114152
-
[32]
G. Consolo, C. Curr` o, G. Grif` o, and G. Valenti. Oscillatory periodic pattern dynamics in hyperbolic reaction-advection-diffusion models. Physical Review E, 105:034206, 2022. doi: 10.1103/PhysRevE.105.034206
-
[33]
G. Consolo, G. Grif` o, and G. Valenti. Dryland vegetation pattern dynamics driven by inertial effects and secondary seed dispersal. Ecological Modelling, 474:110171, 2022. doi: 10.1016/j.ecolmodel.2022.110171
-
[34]
G. Grif` o. Vegetation patterns in the hyperbolic Klausmeier model with secondary seed dispersal. Mathematics, 11:1084, 2023. doi: 10.3390/math11051084
-
[35]
S. Thompson and G. Katul. Secondary seed dispersal and its role in landscape organi- zation. Geophysical Research Letters, 36(2):L02402, 2009. doi: 10.1029/2008GL036044
-
[36]
S. Thompson, S. Assouline, L. Chen, A. Trahktenbrot, T. Svoray, and G. Katul. Sec- ondary dispersal driven by overland flow in drylands: Review and mechanistic model development. Movement Ecology, 2:7, 2014. doi: 10.1186/2051-3933-2-7
-
[37]
A. Marasco, A. Iuorio, F. Carten` ı, G. Bonanomi, F. Giannino, and S Maz- zoleni. Water limitation and negative plant-soil feedback explain vegetation patterns along rainfall gradient. Procedia Environmental Sciences , 19:139–147, 2013. doi: 10.1016/j.proenv.2013.06.016
-
[38]
A. Marasco, A. Iuorio, F. Carten` ı, G. Bonanomi, D.M. Tartakovsky, S. Mazzoleni, and F. Giannino. Vegetation pattern formation due to interactions between water availability and toxicity in plant-soil feedback. Bulletin of Mathematical Biology, 76:2866–2883, 2014. doi: 10.1007/s11538-014-0036-6
-
[39]
A. Iuorio and F. Veerman. The influence of autotoxicity on the dynamics of vegetation spots. Physica D, 427:133015, 2021. doi: 10.1016/j.physd.2021.133015
-
[40]
G. Consolo, G. Grif` o, and G. Valenti. Modeling vegetation patterning on sloped terrains: The role of toxic compounds. Physica D , 459:134020, 2024. doi: 10.1016/j.physd.2023.134020
-
[41]
R. Bastiaansen, P. Carter, and A. Doelman. Stable planar vegetation stripe patterns on sloped terrain in dryland ecosystems. Nonlinearity, 32(8):2759, 2019. doi: 10.1088/1361- 6544/ab1767. 24
-
[42]
L. Eigentler. Species coexistence in resource-limited patterned ecosystems is facilitated by the interplay of spatial self-organisation and intraspecific competition. Oikos, 130: 609–623, 2021. doi: 10.1111/oik.07880
-
[43]
G. Consolo and G. Grif` o. Turing vegetation patterns in flat arid environments with finite soil carrying capacity. Ricerche di Matematica, 2023. doi: 10.1007/s11587-023-00783-z
-
[44]
E. Byrnes, P. Carter, A. Doelman, and L. Liu. Large amplitude radially symmetric spots and gaps in a dryland ecosystem model. Preprint, 2023. doi: 10.48550/arXiv.2208.13167
-
[45]
D.J. Tongway. Banded Vegetation Patterning in Arid and Semiarid Environments . Springer, New York, first edition, 2001. doi: 10.1007/978-1-4613-0207-0
-
[46]
D. Dunkerley. Banded vegetation in some australian semi-arid landscapes: 20 years of field observations to support the development and evaluation of numerical models of vegetation pattern evolution. Desert, 23(2):165–187, 2018. URL https://jdesert.ut. ac.ir/article_69115.html
work page 2018
-
[47]
P. Carter and A. Doelman. Traveling stripes in the Klausmeier model of vegetation pattern formation. SIAM Journal of Applied Mathematics , 78:3213–3237, 2018. doi: 10.1137/18M1196996
-
[48]
L. Sewalt and A. Doelman. Spatially periodic multipulse patterns in a generalized Klaus- meier–Gray–Scott model. SIAM Journal on Applied Dynamical Systems , 16(2):1113– 1163, 2017. doi: 10.1137/16M1078756
-
[49]
J.D. Murray. Mathematical Biology II: Spatial Models and Biomedical Applications . Springer, Berlin, third edition, 2003. doi: 10.1007/b98869
-
[50]
R. Hoyle. Pattern formation. An introduction to methods . Cambridge University Press, New York, first edition, 2007. doi: 10.1017/CBO9780511616051
-
[51]
A. Doelman. Pattern formation in reaction-diffusion systems - an explicit approach. In M.A. Peletier, R.A. van Santen, and E. Siteur, editors, Complexity Science, an Introduc- tion, chapter 4, pages 129–182. World Scientific, 2018. doi: 10.1142/9789813239609 0004
-
[52]
Matlab version: 9.14.0 (r2023a), 2023
The MathWorks Inc. Matlab version: 9.14.0 (r2023a), 2023. URL https://www. mathworks.com
work page 2023
-
[53]
F. Veerman and A. Doelman. Pulses in a Gierer-Meinhardt equation with a slow nonlin- earity. SIAM Journal on Applied Dynamical Systems, 12(1):28–60, 2013. ISSN 1536-0040. doi: 10.1137/120878574. URL http://dx.doi.org/10.1137/120878574
-
[54]
A. Doelman and F. Veerman. An explicit theory for pulses in two component, singularly perturbed, reaction-diffusion equations. Journal of Dynamics and Differential Equations , 27(3):555–595, 2015. ISSN 1040-7294. doi: 10.1007/s10884-013-9325-2. URL http: //dx.doi.org/10.1007/s10884-013-9325-2
-
[55]
A. Doelman, R.A. Gardner, and T.J. Kaper. Large stable pulse solutions in reaction- diffusion equations. Indiana University Mathematics Journal , 50(1):443–507, 2001. ISSN 0022-2518. doi: 10.1512/iumj.2001.50.1873. URL http://dx.doi.org/10.1512/iumj. 2001.50.1873. [link]. 25
-
[56]
E.J. Doedel. AUTO: A program for the automatic bifurcation analysis of autonomous systems. Congressus Numererantium, 30(265-284):25–93, 1981
work page 1981
-
[57]
Nonlinear programming in complex space: Sufficient conditions and duality
N. Fenichel. Geometric singular perturbation theory for ordinary differential equa- tions. J. Differential Equations , 31(1):53–98, 1979. ISSN 0022-0396. doi: 10.1016/0022- 0396(79)90152-9
-
[58]
C.K.R.T. Jones, T.J. Kaper, and N. Kopell. Tracking invariant manifolds up to exponentially small errors. SIAM J. Math. Anal. , 27(2):558–577, 1996. doi: 10.1137/s003614109325966x
-
[59]
C. Kuehn. Multiple Time Scale Dynamics . Springer International Publishing, 2015. doi: 10.1007/978-3-319-12316-5. URL https://doi.org/10.1007%2F978-3-319-12316-5 . 26
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.