Stochastic dynamics at the back of a gene drive propagation wave
Pith reviewed 2026-05-23 01:58 UTC · model grok-4.3
The pith
The probability of wild-type recolonisation after gene drive waves increases with lower drive fitness and smaller carrying capacity.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Gene drive alleles bias their own inheritance and can fix despite fitness costs, but wild-type recolonisation may prevent eradication. We examine conditions for low chance of wild-type recolonisation in one dimension by ensuring the last wild-type individual is surrounded by many drive homozygotes. We make a deterministic approximation of the drive allele distribution within the wave and split the wild-type distribution into deterministic and stochastic parts. Analytical and numerical results indicate that recolonisation probability increases with lower drive fitness and smaller carrying capacity. Simulations confirm extension to two dimensions, with migration rate having smaller impact, and
What carries the argument
Splitting the wild-type allele distribution into deterministic and stochastic components after deterministically approximating the drive allele distribution in the propagation wave.
If this is right
- Recolonisation events are more probable when drive individuals have lower fitness.
- Smaller local carrying capacity raises the chance of wild-type recolonisation.
- The migration rate has a lower impact on recolonisation probability.
- After recolonisation, the chance of drive reinvasion is lower when the population's intrinsic growth rate is smaller.
- These trends hold in two spatial dimensions based on numerical simulations.
Where Pith is reading between the lines
- Gene drive strategies could be adjusted by considering population density to minimize recolonisation risks.
- The decomposition method may help analyze stochastic effects in other ecological traveling waves.
- Field experiments varying carrying capacity could test the predicted dependence on local population size.
Load-bearing premise
The deterministic approximation of the distribution of drive alleles within the wave is sufficiently accurate to permit splitting the wild-type distribution into deterministic and stochastic components.
What would settle it
If simulations or observations show that recolonisation probability does not increase with decreasing drive fitness or decreasing carrying capacity, the central claim would be falsified.
Figures














read the original abstract
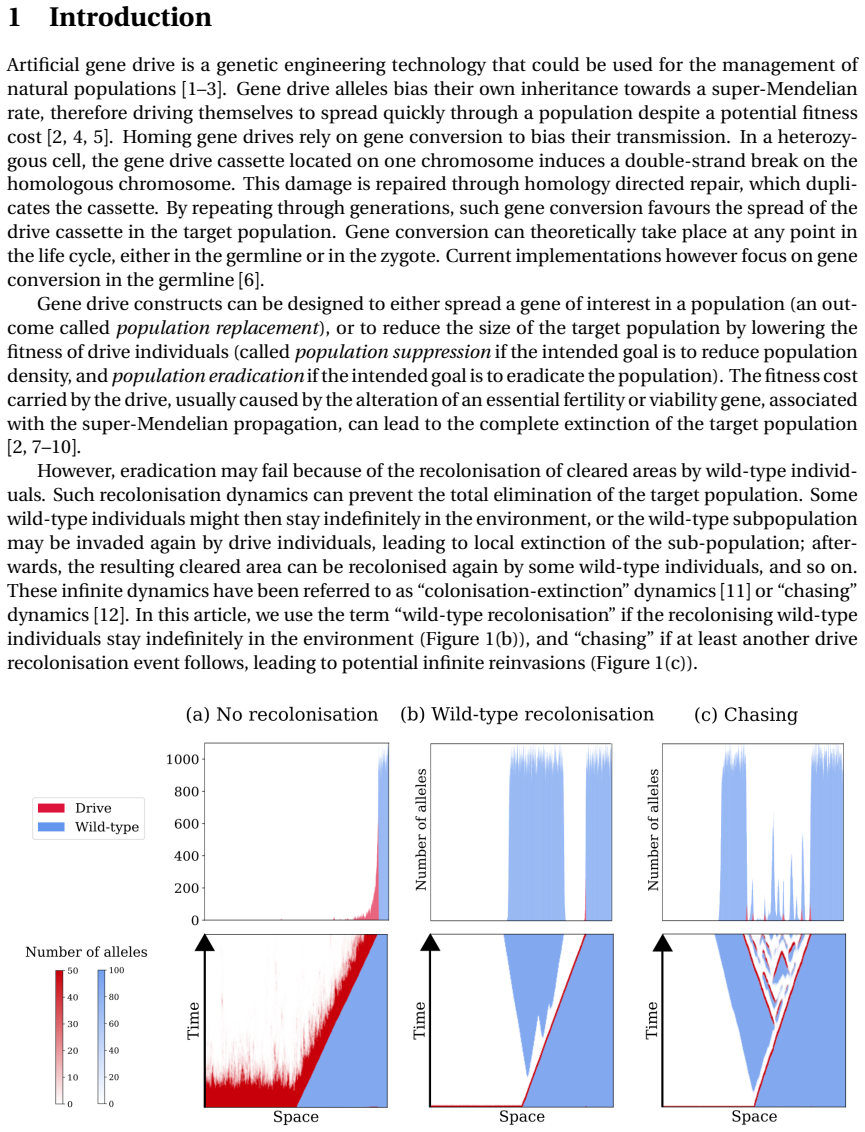
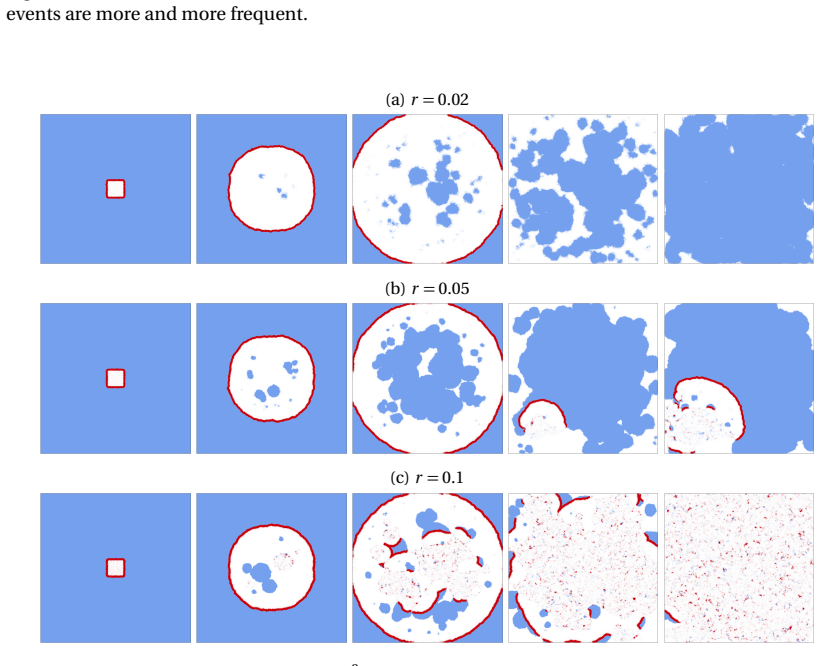
Gene drive alleles bias their own inheritance to offspring. They can fix in a wild-type population in spite of a fitness cost, and even lead to the eradication of the target population if the fitness cost is high. However, this outcome may be prevented or delayed if areas previously cleared by the drive are recolonised by wild-type individuals. Here, we investigate the conditions under which these stochastic wild-type recolonisation events are likely and when they are unlikely to occur in one spatial dimension. More precisely, we examine the conditions ensuring that the last individual carrying a wild-type allele is surrounded by a large enough number of drive homozygous individuals, resulting in a very low chance of wild-type recolonisation. To do so, we make a deterministic approximation of the distribution of drive alleles within the wave, and we split the distribution of wild-type alleles into a deterministic part and a stochastic part. Our analytical and numerical results suggest that the probability of wild-type recolonisation events increases with lower fitness of drive individuals and with smaller local carrying capacity. Numerical simulations show that these results extend to two spatial dimensions. The role of the migration rate however, is less clear but has a lower impact. We further demonstrate that, in the event of wild-type recolonization, the probability of subsequent drive reinvasion decreases with smaller values of the intrinsic growth rate of the population. Overall, our study paves the way for further analysis of wild-type recolonisation at the back of eradication traveling waves.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper claims that a deterministic approximation of the drive allele distribution across a spatial propagation wave, combined with a decomposition of the wild-type allele distribution into deterministic and stochastic components, allows analytical and numerical computation of wild-type recolonization probability behind the wave front. Results indicate this probability rises with lower drive fitness and smaller local carrying capacity; the approach is extended via simulations to two dimensions, with additional analysis of drive reinvasion probability after recolonization.
Significance. If the approximation and decomposition are accurate, the work supplies a useful framework for predicting when gene-drive eradication waves are robust to stochastic recolonization in spatial settings, with direct relevance to the design of drive releases for vector or pest control. The explicit dependence on fitness cost and carrying capacity, together with the 1D-to-2D extension, offers testable predictions that could guide both modeling and field considerations.
major comments (3)
- [Methods / Results (deterministic drive approximation)] The central decomposition (described in the methods and results sections) treats the drive profile as deterministic even in the low-density tail where recolonization is decided. No direct comparison or error quantification against a fully stochastic individual-based model is supplied for this region, leaving the boundary condition supplied to the wild-type stochastic component unvalidated precisely where demographic fluctuations of the drive are expected to matter.
- [Results (dependence on fitness and carrying capacity)] The reported dependence of recolonization probability on drive fitness cost and carrying capacity is obtained from the deterministic-drive plus stochastic-wild-type split. Without sensitivity checks that reintroduce drive stochasticity (or at least quantify its effect on the tail profile), it is unclear whether the claimed trends survive when the weakest assumption is relaxed.
- [Numerical simulations (2D extension)] Abstract and main text state that numerical simulations confirm the 1D trends in 2D, yet no quantitative error bars, convergence tests with respect to lattice size or stochastic realizations, or direct comparison of the analytic approximation to the same 2D stochastic realizations are presented.
minor comments (2)
- [Methods] Notation for the deterministic versus stochastic components of the wild-type density should be introduced with a single consistent symbol set and referenced explicitly when the recolonization probability is derived.
- [Results] The manuscript would benefit from a short table or figure panel that directly juxtaposes the deterministic drive profile against a few stochastic realizations in the tail region, even if only for illustration.
Simulated Author's Rebuttal
We thank the referee for their constructive comments on our manuscript. We address each major point below, indicating where we will revise the paper to incorporate additional validation and details.
read point-by-point responses
-
Referee: The central decomposition (described in the methods and results sections) treats the drive profile as deterministic even in the low-density tail where recolonization is decided. No direct comparison or error quantification against a fully stochastic individual-based model is supplied for this region, leaving the boundary condition supplied to the wild-type stochastic component unvalidated precisely where demographic fluctuations of the drive are expected to matter.
Authors: We acknowledge that the deterministic approximation for the drive profile in the low-density tail is a central modeling choice whose accuracy directly affects the boundary condition for the stochastic wild-type component. While the approximation is motivated by the higher densities in the wave core, we agree that explicit validation in the tail is warranted. In the revised manuscript we will add a direct comparison of the deterministic drive profile against fully stochastic individual-based realizations in the tail region, together with quantitative error measures. revision: yes
-
Referee: The reported dependence of recolonization probability on drive fitness cost and carrying capacity is obtained from the deterministic-drive plus stochastic-wild-type split. Without sensitivity checks that reintroduce drive stochasticity (or at least quantify its effect on the tail profile), it is unclear whether the claimed trends survive when the weakest assumption is relaxed.
Authors: The reported trends follow from the stated decomposition. To assess robustness, we will perform additional individual-based simulations that retain stochasticity in the drive allele and compare the resulting recolonization probabilities (and their dependence on fitness cost and carrying capacity) with the predictions of the decomposed model. Any discrepancies will be quantified and discussed. revision: yes
-
Referee: Abstract and main text state that numerical simulations confirm the 1D trends in 2D, yet no quantitative error bars, convergence tests with respect to lattice size or stochastic realizations, or direct comparison of the analytic approximation to the same 2D stochastic realizations are presented.
Authors: We agree that the 2D simulation section would be strengthened by additional quantitative information. In the revision we will report error bars obtained from multiple independent realizations, present convergence tests with respect to lattice size and number of realizations, and include a direct quantitative comparison between the 1D analytic predictions and the outcomes of the corresponding 2D stochastic simulations. revision: yes
Circularity Check
No significant circularity; derivation is self-contained via independent approximation and simulation
full rationale
The paper's core approach uses a deterministic approximation of drive allele distribution within the wave to decompose the wild-type distribution into deterministic and stochastic parts, then derives recolonization probabilities analytically and numerically from the resulting model. This split and the subsequent claims about dependence on fitness cost and carrying capacity follow directly from the stated equations and simulations without any parameter fitting to the target probability, self-definitional loops, or load-bearing self-citations that reduce the result to its own inputs. The method is externally falsifiable via the described numerical simulations and does not rename known results or import uniqueness via author citations.
Axiom & Free-Parameter Ledger
free parameters (2)
- drive fitness cost
- local carrying capacity
axioms (1)
- domain assumption Deterministic approximation of drive allele distribution within the wave is valid
Reference graph
Works this paper leans on
-
[1]
Nicolas O. Rode et al. “Population management using gene drive: molecular design, models of spread dynamics and assessment of ecological risks”. Conservation Genetics 20.4 (Aug. 1, 2019), pp. 671–690. ISSN : 1572-9737. DOI : 10.1007/s10592-019-01165-5
-
[2]
Austin Burt. “Site-specific selfish genes as tools for the control and genetic engineering of natu- ral populations.”Proceedings of the Royal Society B: Biological Sciences 270.1518 (May 7, 2003), pp. 921–928. ISSN : 0962-8452. DOI : 10.1098/rspb.2002.2319
-
[3]
New developments in the field of genomic technologies and their relevance to conservation management
Gernot Segelbacher et al. “New developments in the field of genomic technologies and their relevance to conservation management”.Conservation Genetics 23.2 (Apr. 1, 2022), pp. 217–242. ISSN : 1572-9737. DOI : 10.1007/s10592-021-01415-5
-
[4]
Standardizing the definition of gene drive
Luke S. Alphey et al. “Standardizing the definition of gene drive”.Proceedings of the National Academy of Sciences 117.49 (Dec. 8, 2020). Publisher: Proceedings of the National Academy of Sciences, pp. 30864– 30867. DOI : 10.1073/pnas.2020417117
-
[5]
Gene Drive: Evolved and Synthetic
Austin Burt and Andrea Crisanti. “Gene Drive: Evolved and Synthetic”. ACS Chemical Biology 13.2 (Feb. 16, 2018). Publisher: American Chemical Society, pp. 343–346. ISSN : 1554-8929. DOI : 10.1021/acschembio.7b01031
-
[6]
Ethan Bier. “Gene drives gaining speed”. Nature Reviews Genetics 23.1 (2022), pp. 5–22
work page 2022
-
[7]
Kyros Kyrou et al. “A CRISPR–Cas9 gene drive targeting doublesex causes complete popula- tion suppression in caged Anopheles gambiae mosquitoes”. Nature Biotechnology 36.11 (Nov. 2018). Number: 11 Publisher: Nature Publishing Group, pp. 1062–1066. ISSN : 1546-1696. DOI : 10.1038/nbt.4245
-
[8]
Gene-drive suppression of mosquito populations in large cages as a bridge between lab and field
Andrew Hammond et al. “Gene-drive suppression of mosquito populations in large cages as a bridge between lab and field”. Nature Communications 12.1 (July 28, 2021). Number: 1 Pub- lisher: Nature Publishing Group, p. 4589.ISSN : 2041-1723. DOI : 10.1038/s41467-021-24790- 6
-
[9]
How driving endonuclease genes can be used to combat pests and disease vectors
H. Charles J. Godfray, Ace North, and Austin Burt. “How driving endonuclease genes can be used to combat pests and disease vectors”. BMC Biology 15.1 (Sept. 11, 2017), p. 81. ISSN : 1741-7007. DOI : 10.1186/s12915-017-0420-4
-
[10]
Demographic feedbacks can hamper the spatial spread of a gene drive
Léo Girardin and Florence Débarre. “Demographic feedbacks can hamper the spatial spread of a gene drive”. Journal of Mathematical Biology 83.6 (Dec. 4, 2021), p. 67. ISSN : 1432-1416. DOI : 10.1007/s00285-021-01702-2
-
[11]
Ace R. North, Austin Burt, and H. Charles J. Godfray. “Modelling the suppression of a malaria vec- tor using a CRISPR-Cas9 gene drive to reduce female fertility”. BMC Biology 18 (Aug. 11, 2020), p. 98. ISSN : 1741-7007. DOI : 10.1186/s12915-020-00834-z
-
[12]
Jackson Champer et al. “Suppression gene drive in continuous space can result in unstable per- sistence of both drive and wild-type alleles”.Molecular Ecology 30.4 (Feb. 2021), pp. 1086–1101. ISSN : 1365-294X. DOI : 10.1111/mec.15788
-
[13]
Gene-drive-mediated extinction is thwarted by population structure and evolution of sib mating
James J Bull, Christopher H Remien, and Stephen M Krone. “Gene-drive-mediated extinction is thwarted by population structure and evolution of sib mating”.Evolution, Medicine, and Public Health 2019.1 (Jan. 1, 2019), pp. 66–81. ISSN : 2050-6201. DOI : 10.1093/emph/eoz014
-
[14]
Modelling the potential of genetic control of malaria mosquitoes at national scale
Ace R. North, Austin Burt, and H. Charles J. Godfray. “Modelling the potential of genetic control of malaria mosquitoes at national scale”. BMC Biology 17.1 (Mar. 29, 2019), p. 26. ISSN : 1741-
work page 2019
-
[15]
DOI : 10.1186/s12915-019-0645-5
-
[16]
Aysegul Birand et al. “Gene drives for vertebrate pest control: Realistic spatial modelling of eradi- cation probabilities and times for island mouse populations”.Molecular Ecology 31.6 (Mar. 2022), pp. 1907–1923. ISSN : 0962-1083. DOI : 10.1111/mec.16361. 28
-
[17]
Samuel E Champer et al. “Anopheles homing suppression drive candidates exhibit unexpected performance differences in simulations with spatial structure”. eLife 11 (Oct. 14, 2022). Ed. by George H Perry, Sebald ANR Verkuijl, and Jim Bull. Publisher: eLife Sciences Publications, Ltd, e79121. ISSN : 2050-084X. DOI : 10.7554/eLife.79121
-
[18]
Yiran Liu et al. “Adversarial interspecies relationships facilitate population suppression by gene drive in spatially explicit models”. Ecology Letters 26.7 (2023), pp. 1174–1185. ISSN : 1461-0248. DOI : 10.1111/ele.14232
-
[19]
Modelling homing suppression gene drive in haplodiploid or- ganisms
Yiran Liu and Jackson Champer. “Modelling homing suppression gene drive in haplodiploid or- ganisms”. Proceedings of the Royal Society B: Biological Sciences 289.1972 (Apr. 13, 2022). Pub- lisher: Royal Society, p. 20220320.DOI : 10.1098/rspb.2022.0320
-
[20]
Jeff F . Paril and Ben L. Phillips. “Slow and steady wins the race: spatial and stochastic processes and the failure of suppression gene drives”. Molecular Ecology (July 5, 2022), mec.16598. ISSN : 0962-1083, 1365-294X. DOI : 10.1111/mec.16598
-
[21]
Yutong Zhu and Jackson Champer. “Simulations Reveal High Efficiency and Confinement of a Population Suppression CRISPR Toxin-Antidote Gene Drive”.ACS Synthetic Biology 12.3 (Mar. 17, 2023). Publisher: American Chemical Society, pp. 809–819.DOI : 10.1021/acssynbio.2c00611
-
[22]
Léna Kläy et al. “Pulled, pushed or failed: the demographic impact of a gene drive can change the nature of its spatial spread”.Journal of Mathematical Biology 87.2 (July 16, 2023), p. 30.ISSN : 1432-1416. DOI : 10.1007/s00285-023-01926-4
-
[23]
James D Murray.Mathematical biology: I. An introduction. Vol. 17. Springer Science & Business Media, 2007
work page 2007
-
[24]
Available: https://link.aps.org/doi/10.1103/PhysRevE
Eric Brunet and Bernard Derrida. “Shift in the velocity of a front due to a cutoff”.Physical Review E 56.3 (Sept. 1, 1997). Publisher: American Physical Society, pp. 2597–2604.DOI : 10.1103/PhysRevE. 56.2597
-
[25]
Effect of Microscopic Noise on Front Propagation
Éric Brunet and Bernard Derrida. “Effect of Microscopic Noise on Front Propagation”.Journal of Statistical Physics 103.1 (Apr. 1, 2001), pp. 269–282. ISSN : 1572-9613. DOI : 10.1023/A:1004875804376
-
[26]
Brunet-Derrida Behavior of Branching-Selection Parti- cle Systems on the Line
Jean Bérard and Jean-Baptiste Gouéré. “Brunet-Derrida Behavior of Branching-Selection Parti- cle Systems on the Line”.Communications in Mathematical Physics 298.2 (Sept. 1, 2010), pp. 323–
work page 2010
-
[27]
DOI : 10.1007/s00220-010-1067-y
ISSN : 1432-0916. DOI : 10.1007/s00220-010-1067-y
-
[28]
Effect of noise on front propagation in reaction- diffusion equations of KPP type
Carl Mueller, Leonid Mytnik, and Jeremy Quastel. “Effect of noise on front propagation in reaction- diffusion equations of KPP type”. Inventiones mathematicae 184.2 (May 1, 2011), pp. 405–453. ISSN : 1432-1297. DOI : 10.1007/s00222-010-0292-5
-
[29]
Recent results on branching random walks
Daniela Bertacchi and Fabio Zucca.Recent results on branching random walks. May 4, 2018.DOI : 10.48550/arXiv.1104.5085. arXiv: 1104.5085[math]
work page internal anchor Pith review Pith/arXiv arXiv doi:10.48550/arxiv.1104.5085 2018
-
[30]
The Wave of Advance of Advantageous Genes
R. A. Fisher. “The Wave of Advance of Advantageous Genes”.Annals of Eugenics 7.4 (1937). _eprint: https://onlinelibrary.wiley.com/doi/pdf/10.1111/j.1469-1809.1937.tb02153.x, pp. 355–369.ISSN : 2050-1439. DOI : 10.1111/j.1469-1809.1937.tb02153.x
-
[31]
Advances in Measuring the Environmental and Social Impacts of Environmental Programs
Jason W . Chapman, V . Alistair Drake, and Don R. Reynolds. “Recent Insights from Radar Studies of Insect Flight”.Annual Review of Entomology 56.1 (2011). _eprint: https://doi.org/10.1146/annurev- ento-120709-144820, pp. 337–356. DOI : 10.1146/annurev-ento-120709-144820
-
[32]
Mass seasonal bioflows of high-flying insect migrants
Gao Hu et al. “Mass seasonal bioflows of high-flying insect migrants”.Science 354.6319 (Dec. 23, 2016). Publisher: American Association for the Advancement of Science, pp. 1584–1587. DOI : 10.1126/science.aah4379
-
[33]
Windborne long-distance migration of malaria mosquitoes in the Sahel
Diana L. Huestis et al. “Windborne long-distance migration of malaria mosquitoes in the Sahel”. Nature 574.7778 (Oct. 2019). Number: 7778 Publisher: Nature Publishing Group, pp. 404–408. ISSN : 1476-4687. DOI : 10.1038/s41586-019-1622-4
-
[34]
Andrea Egizi et al. “The hitchhiker’ s guide to becoming invasive: exotic mosquitoes spread across a US state by human transport not autonomous flight”.Molecular Ecology 25.13 (2016). _eprint: https://onlinelibrary.wiley.com/doi/pdf/10.1111/mec.13653, pp. 3033–3047. ISSN : 1365-294X. DOI : 10.1111/mec.13653
-
[35]
Direct Evidence of Adult Aedes albopictus Dispersal by Car
Roger Eritja et al. “Direct Evidence of Adult Aedes albopictus Dispersal by Car”.Scientific Reports 7.1 (Oct. 24, 2017), p. 14399. ISSN : 2045-2322. DOI : 10.1038/s41598-017-12652-5
-
[36]
Gene drive for population genetic control: non-functional resistance and parental effects
Andrea K. Beaghton et al. “Gene drive for population genetic control: non-functional resistance and parental effects”. Proceedings of the Royal Society B: Biological Sciences 286.1914 (Oct. 30, 2019). Publisher: Royal Society, p. 20191586. DOI : 10.1098/rspb.2019.1586. 29
-
[37]
Andrew M. Hammond et al. “The creation and selection of mutations resistant to a gene drive over multiple generations in the malaria mosquito”. PLOS Genetics 13.10 (Oct. 4, 2017). Pub- lisher: Public Library of Science, e1007039. ISSN : 1553-7404. DOI : 10.1371/journal.pgen. 1007039
-
[38]
Resistance to natural and synthetic gene drive systems
Tom A. R. Price et al. “Resistance to natural and synthetic gene drive systems”.Journal of Evolutionary Biology 33.10 (Oct. 1, 2020), pp. 1345–1360. ISSN : 1010-061X. DOI : 10.1111/jeb.13693. 30
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.