Population dynamics of generalist/specialist strategies in the feast-famine cycles
Pith reviewed 2026-05-22 15:22 UTC · model grok-4.3
The pith
The relative balance between growth and death rates determines whether generalist or specialist strategies dominate microbial populations in fluctuating nutrient environments.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
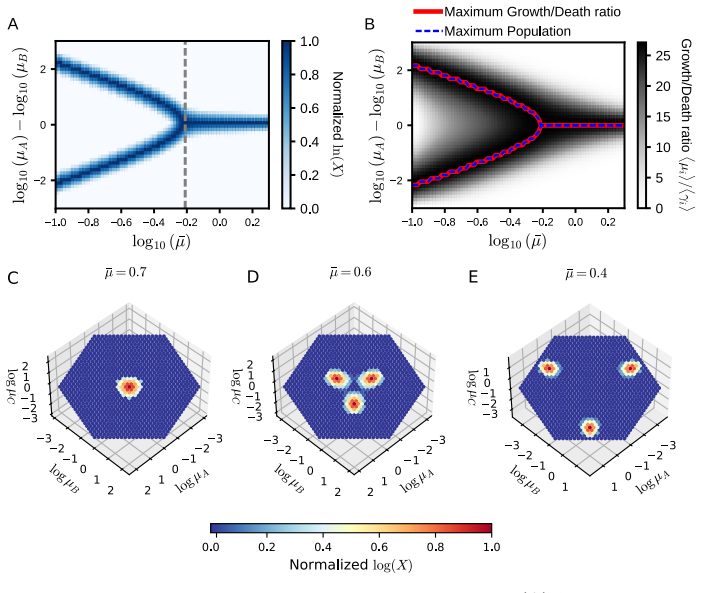
The authors introduce a unified mathematical model that simultaneously incorporates the resource-use trade-off and the growth-death trade-off arising from temporally fluctuating nutrients modeled as discrete stochastic events. They show that the relative balance between growth and death rates critically influences strategy dominance: high average growth rates and weak trade-offs favor generalists, while intense growth-death trade-offs promote specialists.
What carries the argument
Unified mathematical model combining resource-use trade-off with growth-death trade-off under stochastic discrete nutrient supply events.
Load-bearing premise
The growth-death trade-off intersects with the resource-use trade-off in a manner that produces a switch in strategy dominance.
What would settle it
An experiment that varies the strength of the growth-death trade-off while keeping nutrient supply stochastic and observes if specialists take over when the trade-off intensifies.
Figures





read the original abstract
Microbial populations exhibit a broad spectrum of nutrient utilization strategies, ranging from strategies utilizing diverse nutrients, called "generalists," to those being highly adapted to specific nutrients, called "specialists." The mathematical conditions for the diversification of nutrient utilization strategies are central questions in theoretical ecology. Previous studies have shown that trade-offs among different resource utilization functions that cells cannot utilize broad types of substrates at near-maximum speed are crucial for the emergence of diverse strategies. However, in natural settings, nutrient availability often fluctuates over time, imposing additional trade-offs on cells. Cells that grow rapidly under nutrient-rich conditions will suffer a higher death rate under nutrient-poor conditions, creating a growth-death trade-off that intersects with the classical resource-use trade-off. Here, we introduce a unified mathematical model that simultaneously incorporates the resource-use trade-off and the growth-death trade-off. The nutrient supply was modeled as discrete stochastic events, capturing realistic temporal fluctuations. We show that the relative balance between growth and death rates critically influences the dominance of either generalist or specialist strategies. Specifically, under conditions of high average growth rates among different environments and a weak trade-off between growth and death rates, generalists prevail. In contrast, when the growth-death trade-off is intense, specialists emerge as the dominant strategy. Our findings reveal that accounting for the growth-death trade-off is crucial for understanding how microbial communities adapt and evolve in temporally varying environments.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper develops a unified mathematical model for microbial nutrient utilization strategies that integrates classical resource-use trade-offs with an additional growth-death trade-off induced by temporally fluctuating nutrient availability, modeled as discrete stochastic supply events. The central claim is that the relative balance between growth and death rates governs strategy dominance: generalists prevail under high average growth rates across environments combined with weak growth-death trade-offs, whereas intense growth-death trade-offs favor specialists.
Significance. If the reported dominance switch is robust to modeling choices, the work would meaningfully extend prior theoretical ecology results on resource-use trade-offs by incorporating realistic feast-famine dynamics and their associated mortality costs. This could help explain observed microbial diversity in variable environments. However, the abstract supplies no equations, parameter values, or verification steps, and the reader's assessment notes low soundness and potential circularity in the dependence on 'relative balance' and trade-off intensity, limiting immediate impact.
major comments (2)
- [Model construction (unified mathematical model)] The intersection of the growth-death trade-off with the resource-use trade-off is load-bearing for the central claim yet under-specified. The abstract states that 'the growth-death trade-off is assumed to intersect with the resource-use trade-off in a form that produces the reported dominance switch,' but provides no explicit functional form (linear, hyperbolic, or otherwise) linking maximum growth rate to famine death rate, nor any sensitivity analysis to alternative shapes or stochastic event frequencies.
- [Results and discussion] The reported conditions for generalist dominance ('high average growth rates among different environments and a weak trade-off') versus specialist dominance ('intense' trade-off) appear to hinge on the specific parameterization of the discrete stochastic nutrient supply process. Without reported robustness checks or explicit equations, it is unclear whether the switch is a general outcome or an artifact of fixed event magnitude/frequency choices.
minor comments (2)
- [Abstract] The abstract would be strengthened by including at least one key model equation or a brief statement of the trade-off functional form to allow readers to assess the claimed conditions without the full text.
- [Introduction] Notation for 'relative balance' and 'intensity' of trade-offs should be defined more precisely when first introduced to avoid ambiguity in interpreting the dominance results.
Simulated Author's Rebuttal
We thank the referee for their detailed review and constructive suggestions. We have carefully considered the comments regarding model specification and robustness of results. We address each point below and plan to incorporate clarifications and additional analyses in the revised manuscript.
read point-by-point responses
-
Referee: [Model construction (unified mathematical model)] The intersection of the growth-death trade-off with the resource-use trade-off is load-bearing for the central claim yet under-specified. The abstract states that 'the growth-death trade-off is assumed to intersect with the resource-use trade-off in a form that produces the reported dominance switch,' but provides no explicit functional form (linear, hyperbolic, or otherwise) linking maximum growth rate to famine death rate, nor any sensitivity analysis to alternative shapes or stochastic event frequencies.
Authors: We agree that the abstract does not provide the explicit functional form or sensitivity analysis. In the full manuscript (Methods section), the growth-death trade-off is implemented by setting the famine death rate proportional to the maximum growth rate via an intensity parameter (linear form d = τ μ_max). We will update the abstract to state this functional form explicitly and add a new subsection with sensitivity analyses for alternative forms (e.g., hyperbolic) and a range of stochastic event frequencies and magnitudes. revision: yes
-
Referee: [Results and discussion] The reported conditions for generalist dominance ('high average growth rates among different environments and a weak trade-off') versus specialist dominance ('intense' trade-off) appear to hinge on the specific parameterization of the discrete stochastic nutrient supply process. Without reported robustness checks or explicit equations, it is unclear whether the switch is a general outcome or an artifact of fixed event magnitude/frequency choices.
Authors: The reported dominance switch is obtained from both analytical approximations of the stochastic process and numerical simulations across parameter regimes, as detailed in the Results. We acknowledge that additional explicit robustness checks would strengthen the claim of generality. In the revision we will add supplementary figures and text showing that the qualitative switch between generalist and specialist dominance persists when event magnitude and frequency are varied over an order of magnitude. revision: yes
Circularity Check
Model exploration of trade-off balance yields strategy dominance without definitional reduction or fitted predictions
full rationale
The paper defines a unified dynamical model that explicitly incorporates both the classical resource-use trade-off and an additional growth-death trade-off, with nutrient supply implemented as discrete stochastic events. It then varies parameters representing average growth rates and the intensity of the growth-death trade-off to observe shifts in dominance between generalist and specialist strategies. No equations or sections reduce the reported outcomes to fitted parameters by construction, nor do they rely on self-citations for uniqueness theorems or ansatzes that would make the central claim tautological. The results are generated from forward simulation or analysis of the independently specified model equations, making the derivation self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
free parameters (1)
- growth-death trade-off intensity
axioms (1)
- domain assumption Nutrient availability fluctuates over time as discrete stochastic events.
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
γ_{i,e}=a·exp(b μ_{i,e}) (Eq.5); r=⟨μ⟩/⟨γ⟩; phenotype β invades when μ_β/γ_β > μ_α/γ_α (Eq.10)
-
IndisputableMonolith/Foundation/BranchSelection.leanbranch_selection unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
strong convex resource-use trade-off (product μ_{i,e})^{1/E}=μ_bar (Eq.4) and stochastic feast-famine supply
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Larsen BB, Miller EC, Rhodes MK, Wiens JJ. Inordinate Fondness Multiplied and Redistributed: The Number of Species on Earth and the New Pie of Life. The Quarterly Review of Biology. 2017 Sep;92(3):229-65. doi:10.1086/693564
-
[2]
Prokaryotes: The Unseen Majority
Whitman WB, Coleman DC, Wiebe WJ. Prokaryotes: The Unseen Majority. Proceedings of the National Academy of Sciences. 1998 Jun;95(12):6578-83. doi:10.1073/pnas.95.12.6578
-
[3]
Will a Large Complex System Be Stable? Nature
May RM. Will a Large Complex System Be Stable? Nature. 1972 Aug;238(5364):413-4. doi:10.1038/238413a0
-
[4]
The Feasibility and Stability of Large Complex Biological Networks: A Random Matrix Approach
Stone L. The Feasibility and Stability of Large Complex Biological Networks: A Random Matrix Approach. Scientific Reports. 2018 May;8(1):8246. doi:10.1038/s41598-018-26486-2
-
[5]
Mechanisms of Maintenance of Species Diversity
Chesson P. Mechanisms of Maintenance of Species Diversity. Annual Review of Ecology and Systematics. 2000 Nov;31(1):343-66. doi:10.1146/annurev.ecolsys.31.1.343
-
[6]
Niche Partitioning Facilitates Coexistence of Closely Related Honey Bee Gut Bacteria
Brochet S, Quinn A, Mars RA, Neuschwander N, Sauer U, Engel P. Niche Partitioning Facilitates Coexistence of Closely Related Honey Bee Gut Bacteria. eLife. 2021 Jul;10:e68583. doi:10.7554/eLife.68583
-
[7]
Yu XA, McLean C, Hehemann JH, Angeles-Albores D, Wu F, Muszy´ nski A, et al. Low-Level Resource Partitioning Supports Coexistence among Functionally Redundant Bacteria during Successional Dynamics. The ISME Journal. 2024 Jan;18(1):wrad013. doi:10.1093/ismejo/wrad013
-
[8]
Trade-Offs in Community Ecology: Linking Spatial Scales and Species Coexistence
Kneitel JM, Chase JM. Trade-Offs in Community Ecology: Linking Spatial Scales and Species Coexistence. Ecology Letters. 2004;7(1):69-80. doi:10.1046/j.1461-0248.2003.00551.x
-
[9]
Ekkers DM, Tusso S, Moreno-Gamez S, Rillo MC, Kuipers OP. Trade-Offs Predicted by Metabolic Network Structure Give Rise to Evolutionary Specialization and Phenotypic Diversification. Molecular Biology and Evolution. 2022 Jun;39(6):msac124. doi:10.1093/molbev/msac124
-
[10]
Kilham SS. Dynamics of Lake Michigan Natural Phytoplankton Communities in Continuous Cultures Along a Si:P Loading Gradient. Canadian Journal of Fisheries and Aquatic Sciences. 1986 Feb;43(2):351-60. doi:10.1139/f86-045
-
[11]
Edwards KF, Klausmeier CA, Litchman E. Evidence for a Three-Way Trade-off between Nitrogen and Phosphorus Competitive Abilities and Cell Size in Phytoplankton. Ecology. 2011;92(11):2085-95. arXiv:23034941
work page 2011
-
[12]
Interdependence of Cell Growth and Gene Expression: Origins and Consequences
Scott M, Gunderson CW, Mateescu EM, Zhang Z, Hwa T. Interdependence of Cell Growth and Gene Expression: Origins and Consequences. Science. 2010 Nov;330(6007):1099-102. doi:10.1126/science.1192588. May 1, 2026 23/26
-
[13]
Quantitative Proteomic Analysis Reveals a Simple Strategy of Global Resource Allocation in Bacteria
Hui S, Silverman JM, Chen SS, Erickson DW, Basan M, Wang J, et al. Quantitative Proteomic Analysis Reveals a Simple Strategy of Global Resource Allocation in Bacteria. Molecular Systems Biology. 2015 Feb;11(1):784. doi:10.15252/msb.20145697
-
[14]
Evolution of Diversity in Metabolic Strategies
Caetano R, Ispolatov Y, Doebeli M. Evolution of Diversity in Metabolic Strategies. eLife. 2021 Aug;10:e67764. doi:10.7554/eLife.67764
-
[15]
Schink SJ, Christodoulou D, Mukherjee A, Athaide E, Brunner V, Fuhrer T, et al. Glycolysis/Gluconeogenesis Specialization in Microbes Is Driven by Biochemical Constraints of Flux Sensing. Molecular Systems Biology. 2022 Jan;18(1):e10704. doi:10.15252/msb.202110704
-
[16]
The Evolution of Ecological Specialization
Futuyma DJ, Moreno G. The Evolution of Ecological Specialization. Annual Review of Ecology and Systematics. 1988;19:207-33. arXiv:2097153
work page 1988
-
[17]
Evolution of Ecological Niche Breadth
Sexton JP, Montiel J, Shay JE, Stephens MR, Slatyer RA. Evolution of Ecological Niche Breadth. Annual Review of Ecology, Evolution, and Systematics. 2017;48(1):183-206. doi:10.1146/annurev-ecolsys-110316-023003
-
[18]
Fluctuation Induces Evolutionary Branching in a Mathematical Model of Ecosystems
Tachikawa M. Fluctuation Induces Evolutionary Branching in a Mathematical Model of Ecosystems. PLOS ONE. 2008 Dec;3(12):e3925. doi:10.1371/journal.pone.0003925
-
[19]
Cooper TF, Lenski RE. Experimental Evolution with E. Coli in Diverse Resource Environments. I. Fluctuating Environments Promote Divergence of Replicate Populations. BMC Evolutionary Biology. 2010 Jan;10(1):11. doi:10.1186/1471-2148-10-11
-
[20]
Coexistence in a Variable Environment: Eco-evolutionary Perspectives
Kremer CT, Klausmeier CA. Coexistence in a Variable Environment: Eco-evolutionary Perspectives. Journal of Theoretical Biology. 2013 Dec;339:14-25. doi:10.1016/j.jtbi.2013.05.005
-
[21]
The Experimental Evolution of Specialists, Generalists, and the Maintenance of Diversity
Kassen R. The Experimental Evolution of Specialists, Generalists, and the Maintenance of Diversity. Journal of Evolutionary Biology. 2002;15(2):173-90. doi:10.1046/j.1420-9101.2002.00377.x
-
[22]
Tuning Environmental Timescales to Evolve and Maintain Generalists
Sachdeva V, Husain K, Sheng J, Wang S, Murugan A. Tuning Environmental Timescales to Evolve and Maintain Generalists. Proceedings of the National Academy of Sciences. 2020 Jun;117(23):12693-9. doi:10.1073/pnas.1914586117
-
[23]
Cycles of Famine and Feast: The Starvation and Outgrowth Strategies of a marineVibrio
Srinivasan S, Kjelleberg S. Cycles of Famine and Feast: The Starvation and Outgrowth Strategies of a marineVibrio. Journal of Biosciences. 1998 Oct;23(4):501-11. doi:10.1007/BF02936144
-
[24]
Microbial Ecology of Organic Aggregates in Aquatic Ecosystems
Simon M, Grossart HP, Schweitzer B, Ploug H. Microbial Ecology of Organic Aggregates in Aquatic Ecosystems. Aquatic Microbial Ecology. 2002 Jun;28(2):175-211. doi:10.3354/ame028175
-
[25]
Frequency- and Amplitude-Dependent Microbial Population Dynamics during Cycles of Feast and Famine
Merritt J, Kuehn S. Frequency- and Amplitude-Dependent Microbial Population Dynamics during Cycles of Feast and Famine. Physical Review Letters. 2018 Aug;121(9):098101. doi:10.1103/PhysRevLett.121.098101
-
[26]
Dynamics of Bacterial Populations under the Feast-Famine Cycles
Himeoka Y, Mitarai N. Dynamics of Bacterial Populations under the Feast-Famine Cycles. Physical Review Research. 2020 Mar;2(1):013372. doi:10.1103/PhysRevResearch.2.013372. May 1, 2026 24/26
-
[27]
Notley-McRobb L, King T, Ferenci T. rpoS Mutations and Loss of General Stress Resistance in Escherichia Coli Populations as a Consequence of Conflict between Competing Stress Responses. Journal of Bacteriology. 2002 Feb;184(3):806-11. doi:10.1128/JB.184.3.806-811.2002
-
[28]
Proteases and Protein Degradation in Escherichia Coli
Maurizi MR. Proteases and Protein Degradation in Escherichia Coli. Experientia. 1992 Feb;48(2):178-201. doi:10.1007/BF01923511
-
[29]
Escherichia Coli Can Survive Stress by Noisy Growth Modulation
Patange O, Schwall C, Jones M, Villava C, Griffith DA, Phillips A, et al. Escherichia Coli Can Survive Stress by Noisy Growth Modulation. Nature Communications. 2018 Dec;9(1):5333. doi:10.1038/s41467-018-07702-z
-
[30]
Biselli E, Schink SJ, Gerland U. Slower Growth of Escherichia Coli Leads to Longer Survival in Carbon Starvation Due to a Decrease in the Maintenance Rate. Molecular Systems Biology. 2020 Jun;16(6):e9478. doi:10.15252/msb.20209478
-
[31]
Evolution Restricts the Coexistence of Specialists and Generalists: The Role of Trade-off Structure
Egas M, Dieckmann U, Sabelis MW. Evolution Restricts the Coexistence of Specialists and Generalists: The Role of Trade-off Structure. The American Naturalist. 2004 Apr;163(4):518-31. doi:10.1086/382599
-
[32]
Trait–Fitness Relationships Determine How Trade-off Shapes Affect Species Coexistence
Ehrlich E, Becks L, Gaedke U. Trait–Fitness Relationships Determine How Trade-off Shapes Affect Species Coexistence. Ecology. 2017;98(12):3188-98. doi:10.1002/ecy.2047
-
[33]
Growth Tradeoffs Produce Complex Microbial Communities on a Single Limiting Resource
Manhart M, Shakhnovich EI. Growth Tradeoffs Produce Complex Microbial Communities on a Single Limiting Resource. Nature Communications. 2018 Aug;9(1):3214. doi:10.1038/s41467-018-05703-6
-
[34]
Nutrient Levels and Trade-Offs Control Diversity in a Serial Dilution Ecosystem
Erez A, Lopez JG, Weiner BG, Meir Y, Wingreen NS. Nutrient Levels and Trade-Offs Control Diversity in a Serial Dilution Ecosystem. eLife. 2020 Sep;9:e57790. doi:10.7554/eLife.57790
-
[35]
Fine-Scale Diversity of Microbial Communities Due to Satellite Niches in Boom and Bust Environments
Fridman Y, Wang Z, Maslov S, Goyal A. Fine-Scale Diversity of Microbial Communities Due to Satellite Niches in Boom and Bust Environments. PLOS Computational Biology. 2022 Dec;18(12):e1010244. doi:10.1371/journal.pcbi.1010244
-
[36]
Diauxic Lags Explain Unexpected Coexistence in Multi-resource Environments
Bloxham B, Lee H, Gore J. Diauxic Lags Explain Unexpected Coexistence in Multi-resource Environments. Molecular Systems Biology. 2022 May;18(5):MSB202110630. doi:10.15252/msb.202110630
-
[37]
Bloxham B, Lee H, Gore J. Biodiversity Is Enhanced by Sequential Resource Utilization and Environmental Fluctuations via Emergent Temporal Niches. PLOS Computational Biology. 2024 May;20(5):e1012049. doi:10.1371/journal.pcbi.1012049
-
[38]
NUTRIENT CYCLING IN MOIST TROPICAL FOREST
Vitousek PM, Jr RLS. NUTRIENT CYCLING IN MOIST TROPICAL FOREST. Annual Review of Ecology, Evolution, and Systematics. 1986 Nov;17(Volume 17,):137-67. doi:10.1146/annurev.es.17.110186.001033
-
[39]
Levins R, Evolution in Changing Environments: Some Theoretical Explorations, Princeton University Press; 1968
work page 1968
-
[40]
Thermodynamic Constraints Shape the Structure of Carbon Fixation Pathways
Bar-Even A, Flamholz A, Noor E, Milo R. Thermodynamic Constraints Shape the Structure of Carbon Fixation Pathways. Biochimica et Biophysica Acta (BBA) - Bioenergetics. 2012 Sep;1817(9):1646-59. doi:10.1016/j.bbabio.2012.05.002. May 1, 2026 25/26
-
[41]
Source–Sink Dynamics Shape the Evolution of Antibiotic Resistance and Its Pleiotropic Fitness Cost
Perron GG, Gonzalez A, Buckling A. Source–Sink Dynamics Shape the Evolution of Antibiotic Resistance and Its Pleiotropic Fitness Cost. Proceedings of the Royal Society B: Biological Sciences. 2007 Sep;274(1623):2351-6. doi:10.1098/rspb.2007.0640
-
[42]
Sources and Sinks: A Stochastic Model of Evolution in Heterogeneous Environments
Hermsen R, Hwa T. Sources and Sinks: A Stochastic Model of Evolution in Heterogeneous Environments. Physical Review Letters. 2010 Dec;105(24):248104. doi:10.1103/PhysRevLett.105.248104
-
[43]
Generalist Species Drive Microbial Dispersion and Evolution
Sriswasdi S, Yang Cc, Iwasaki W. Generalist Species Drive Microbial Dispersion and Evolution. Nature Communications. 2017 Oct;8(1):1162. doi:10.1038/s41467-017-01265-1
-
[44]
A Sink Host Allows a Specialist Herbivore to Persist in a Seasonal Source
Laska A, Magalh˜ aes S, Lewandowski M, Puchalska E, Karpicka-Ignatowska K, Radwa´ nska A, et al. A Sink Host Allows a Specialist Herbivore to Persist in a Seasonal Source. Proceedings of the Royal Society B: Biological Sciences;288(1958):20211604. doi:10.1098/rspb.2021.1604. May 1, 2026 26/26
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.