Beyond water limitation in vegetation-autotoxicity patterning: a cross-diffusion model
Pith reviewed 2026-05-19 10:51 UTC · model grok-4.3
The pith
A biomass-toxicity cross-diffusion model produces stable Turing vegetation patterns without water dynamics.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The central claim is that a cross-diffusion model derived as the fast-reaction limit of a three-species system with biomass, toxicity, and an auxiliary variable supports the formation of stable Turing patterns in vegetation for a wide range of parameter values, thanks to the cross-diffusion term, even without explicit water dynamics.
What carries the argument
The cross-diffusion term in the reduced biomass-toxicity equations, obtained from the fast-reaction limit of the three-species system, which allows toxicity to influence biomass movement and thereby generate the instability required for Turing patterns.
If this is right
- Vegetation patterns can form in the absence of water limitation driven only by autotoxicity feedback.
- The model adds propagation reduction to the previously considered effects of growth inhibition and increased mortality.
- Pattern formation occurs robustly for broad parameter ranges as verified by linear analysis, simulations, and numerical continuation.
- The framework applies to any ecological interaction reducible to a fast-reaction limit with cross-diffusion.
Where Pith is reading between the lines
- The same reduction technique could generate cross-diffusion models for autotoxicity in microbial or algal systems.
- Controlled experiments with uniform moisture but varying toxin levels could test whether patterns appear as predicted.
- Reducing autotoxin production might offer a practical way to suppress unwanted spatial patterning in crops.
- The approach connects to other fast-limit derivations in chemical and physical pattern-forming systems.
Load-bearing premise
The three-species system with separated time scales must admit a well-defined fast-reaction limit that produces exactly the cross-diffusion terms used in the two-equation model.
What would settle it
Numerical simulations of the two-equation model with the cross-diffusion coefficient set to zero should produce no spatial patterns while nonzero values should produce them; equivalently, field data showing stable vegetation patterns in autotoxic systems with uniform water supply would support the claim.
Figures



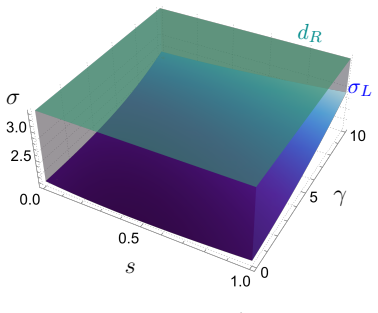
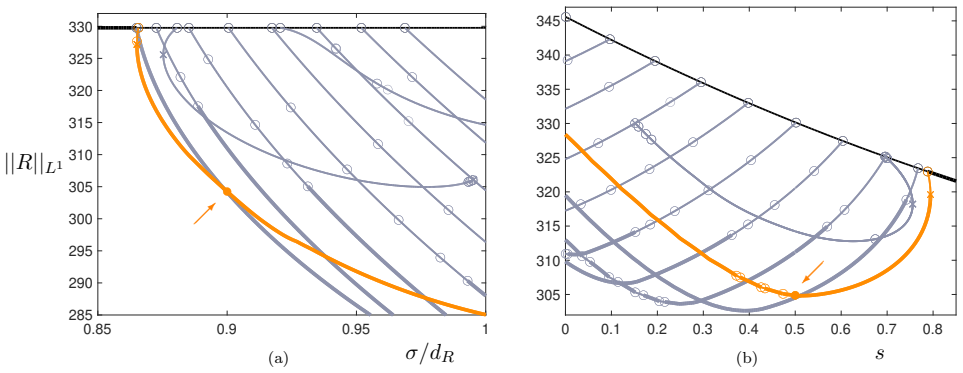
read the original abstract
Many mathematical models describing vegetation patterns are based on biomass--water interactions, due to the impact of this limited resource in arid and semi-arid environments. However, in recent years, a novel biological factor called autotoxicity has proved to play a key role in vegetation spatiotemporal dynamics, particularly by inhibiting biomass growth and increasing its natural mortality rate. In a standard reaction-diffusion framework, biomass-toxicity dynamics alone are unable to support the emergence of stable spatial patterns. In this paper, we derive a cross-diffusion model for biomass and toxicity dynamics as the fast-reaction limit of a three-species system involving dichotomy and different time scales. Within this general framework, in addition to growth inhibition and extra-mortality already considered in previous studies, the additional effect of ''propagation reduction'' induced by autotoxicity on vegetation dynamics is obtained. By combining linearised analysis, simulations, and continuation, we investigate the formation of spatial patterns. Thanks to the cross-diffusion term, for the first time, a spatial model based solely on biomass-toxicity feedback without explicit water dynamics supports the formation of stable (Turing) vegetation patterns for a wide range of parameter values.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper derives a two-equation cross-diffusion system for biomass and toxicity as the fast-reaction limit of a three-species model with dichotomy and separated timescales. It performs linear stability analysis, numerical simulations, and continuation to show that the cross-diffusion terms enable stable Turing patterns over a wide parameter range, providing the first such spatial model based solely on biomass-toxicity feedback without explicit water dynamics.
Significance. If the reduced model is a faithful limit, the work supplies a new mechanism for vegetation patterning driven by autotoxicity alone. The combination of linearised analysis, simulations, and continuation provides concrete evidence for pattern formation across parameter values, which is a methodological strength. The explicit fast-reaction derivation from an independent three-species system (rather than ad-hoc fitting) adds transparency to the modelling assumptions.
major comments (2)
- [Model derivation] Model derivation section: the reduction to the specific cross-diffusion terms in the two-equation system is presented as the fast-reaction limit of the three-species model, but no convergence theorem, uniform estimates, or error bounds are supplied to guarantee that the limit exists and reproduces exactly those terms (including the propagation-reduction effect). This is load-bearing for the central claim, because the Turing analysis and all subsequent simulations are performed on the reduced system; if additional reaction terms survive or the cross terms differ, the reported patterns do not correspond to the stated biological mechanism.
- [Abstract and §4] Abstract and §4 (numerical results): the claim of patterns 'for a wide range of parameter values' is not accompanied by explicit intervals, robustness metrics, or error measures on the continuation or simulation results. Without these, the quantitative support for the 'wide range' assertion remains only moderate.
minor comments (2)
- [Model derivation] Notation for the cross-diffusion coefficients should be introduced with a clear table or explicit definitions immediately after the reduced equations to improve readability.
- [Numerical results] Figure captions for the simulation panels should state the precise parameter values and initial conditions used, rather than referring only to 'typical' values.
Simulated Author's Rebuttal
We thank the referee for the positive evaluation of the significance of our work and for the detailed, constructive comments. We respond to each major comment below.
read point-by-point responses
-
Referee: [Model derivation] Model derivation section: the reduction to the specific cross-diffusion terms in the two-equation system is presented as the fast-reaction limit of the three-species model, but no convergence theorem, uniform estimates, or error bounds are supplied to guarantee that the limit exists and reproduces exactly those terms (including the propagation-reduction effect). This is load-bearing for the central claim, because the Turing analysis and all subsequent simulations are performed on the reduced system; if additional reaction terms survive or the cross terms differ, the reported patterns do not correspond to the stated biological mechanism.
Authors: We acknowledge that the derivation in the manuscript is formal and does not include a rigorous convergence theorem, uniform estimates, or error bounds. The reduction is obtained by the standard singular-perturbation procedure for fast-reaction limits with separated timescales, which produces the stated cross-diffusion terms and the propagation-reduction effect. In the revision we will expand the model-derivation section with an explicit step-by-step asymptotic calculation and add a short discussion of the modelling assumptions under which the formal limit is expected to hold, together with references to analogous reductions in the literature. A complete rigorous convergence analysis lies outside the scope of the present biologically focused study. revision: partial
-
Referee: [Abstract and §4] Abstract and §4 (numerical results): the claim of patterns 'for a wide range of parameter values' is not accompanied by explicit intervals, robustness metrics, or error measures on the continuation or simulation results. Without these, the quantitative support for the 'wide range' assertion remains only moderate.
Authors: We agree that the quantitative support for the 'wide range' claim can be strengthened. In the revised manuscript we will augment §4 with explicit intervals for the key parameters (cross-diffusion coefficients, reaction rates, and diffusion ratios) over which stable Turing patterns are found by numerical continuation. We will also report robustness metrics (e.g., the fraction of sampled parameter space yielding patterns) and include numerical error indicators from the simulations to make the evidence more precise. revision: yes
Circularity Check
Derivation via fast-reaction limit of independent three-species system is self-contained and non-circular
full rationale
The paper derives the two-equation cross-diffusion model explicitly as the fast-reaction limit of a separate three-species system with dichotomy and separated timescales. This limit process supplies the cross terms as an output rather than defining them in terms of the target Turing patterns or fitting them to pattern data. The subsequent linear stability analysis, simulations, and continuation are performed on the derived reduced system; they do not feed back into the derivation step. No self-definitional loop, fitted-input-as-prediction, or load-bearing self-citation chain appears in the model-construction chain. The cited prior work on autotoxicity effects is used only for the reaction terms already studied elsewhere and is not required to justify the cross-diffusion structure itself. The derivation therefore remains independent of the final pattern-formation claims.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption The three-species system with dichotomy and different time scales possesses a well-defined fast-reaction limit yielding the cross-diffusion model.
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
we derive a cross-diffusion model for biomass and toxicity dynamics as the fast-reaction limit of a three-species system involving dichotomy and different time scales
-
IndisputableMonolith/Foundation/AlexanderDuality.leanalexander_duality_circle_linking unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
Thanks to the cross-diffusion term, for the first time, a spatial model based solely on biomass-toxicity feedback without explicit water dynamics supports the formation of stable (Turing) vegetation patterns
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
- [1]
- [2]
-
[3]
M. Allegrezza, G. Bonanomi, M. Zotti, F. Cartenì, M. Moreno, L. Olivieri, M. Garbarino, G. Tesei, F. Giannino, and S. Mazzoleni. Biogeography and shape of fungal fairy rings in the Apennine mountains, Italy.Journal of Biogeography, 49(2):353–363, 2022
work page 2022
-
[4]
R. Bastiaansen, M. Chirilus-Bruckner, and A. Doelman. Pulse solutions for an extended Klaus- meier model with spatially varying coefficients.SIAM Journal on Applied Dynamical Systems, 19(1):1–57, 2020
work page 2020
-
[5]
R. Bastiaansen and A. Doelman. The dynamics of disappearing pulses in a singularly perturbed reaction–diffusion system with parameters that vary in time and space.Physica D: Nonlinear Phenomena, 388:45–72, 2019
work page 2019
-
[6]
R. Bastiaansen, A. Doelman, M.B. Eppinga, and M. Rietkerk. The effect of climate change on the resilience of ecosystems with adaptive spatial pattern formation.Ecology Letters, 23(3):414–429, 2020
work page 2020
-
[7]
B.K. Bera, O. Tzuk, J.J.R. Bennett, U. Dieckmann, and E. Meron. Can spatial self-organization inhibit evolutionary adaptation?Journal of The Royal Society Interface, 22(222):20240454, 2025
work page 2025
-
[8]
G. Bonanomi, G. Incerti, A. Stinca, F. Cartenì, F. Giannino, and S. Mazzoleni. Ring formation in clonal plants.Community Ecology, 15(1):77–86, 2014
work page 2014
-
[9]
G. Bonanomi, M. Zotti, M. Idbella, P. Termolino, V. De Micco, and S. Mazzoleni. Field ev- idence for litter and self-DNA inhibitory effects on Alnus glutinosa roots. New Phytologist, 236(2):399–412, 2022. 14
work page 2022
-
[10]
F. Cartenì, A. Marasco, G. Bonanomi, S. Mazzoleni, M. Rietkerk, and F. Giannino. Negative plant soil feedback explaining ring formation in clonal plants. Journal of Theoretical Biology, 313:153–161, 2012
work page 2012
-
[11]
P. Carter and A. Doelman. Traveling stripes in the Klausmeier model of vegetation pattern formation. SIAM Journal on Applied Mathematics, 78(6):3213–3237, 2018
work page 2018
- [12]
-
[13]
G. Consolo, G. Grifò, and G. Valenti. Modelling vegetation patterning on sloped terrains: The role of toxic compounds.Physica D: Nonlinear Phenomena, 459:134020, 2023
work page 2023
-
[14]
S. De Baets, J. Poesen, A. Knapen, G.G. Barberá, and J.A. Navarro. Root characteristics of rep- resentative Mediterranean plant species and their erosion-reducing potential during concentrated runoff. Plant and Soil, 294(1–2):169–183, 2007
work page 2007
-
[15]
Non-triangularcross-diffusionsystemswithpredator–preyreaction terms
L.DesvillettesandC.Soresina. Non-triangularcross-diffusionsystemswithpredator–preyreaction terms. Ricerche di Matematica, 68:295–314, 2019
work page 2019
- [16]
- [17]
- [18]
-
[19]
D. Fanelli, C. Cianci, and F. Di Patti. Turing instabilities in reaction-diffusion systems with cross diffusion. The European Physical Journal B, 86(4):1–8, 2013
work page 2013
- [20]
- [21]
- [22]
- [23]
-
[24]
M. Iida, M. Mimura, and H. Ninomiya. Diffusion, cross-diffusion and competitive interaction. Journal of Mathematical Biology, 53(4):617–641, 2006
work page 2006
-
[25]
A. Iuorio and F. Veerman. The influence of autotoxicity on the dynamics of vegetation spots. Physica D: Nonlinear Phenomena, 427:133015, 2021
work page 2021
- [26]
-
[27]
S. Kéfi, M. Rietkerk, M. van Baalen, and M. Loreau. Local facilitation, bistability and transitions in arid ecosystems.Theoretical Population Biology, 71(3):367–379, 2007
work page 2007
-
[28]
S. Kéfi, V. Guttal, W.A. Brock, S.R. Carpenter, V. Dakos, et al. Early warning signals of ecological transitions: Methods for spatial patterns.PLoS ONE, 9(3):e92097, 2014
work page 2014
-
[29]
C.A. Klausmeier. Regular and irregular patterns in semiarid vegetation.Science, 284(5421):1826– 1828, 1999
work page 1999
-
[30]
C. Kuehn and C. Soresina. Numerical continuation for a fast-reaction system and its cross-diffusion limit. SN Partial Differential Equations and Applications, 1(2):1–26, 2020
work page 2020
-
[31]
A. Marasco, F. Giannino, and A. Iuorio. Modelling competitive interactions and plant–soil feed- back in vegetation dynamics.Ricerche di Matematica, 69(2):553–577, 2020
work page 2020
-
[32]
A. Marasco, A. Iuorio, F. Cartenì, G. Bonanomi, D.M. Tartakovsky, S. Mazzoleni, and F. Gian- nino. Vegetation pattern formation due to interactions between water availability and toxicity in plant-soil feedback.Bulletin of Mathematical Biology, 76(11):2866–2883, 2014
work page 2014
-
[33]
F. Martínez, O. Merino, A. Martín, D. García Martín, and J. Merino. Belowground structure and production in a mediterranean sand dune shrub community.Plant and Soil, 201:209–216, 1998
work page 1998
-
[34]
R. Martínez-García, C. Cabal, J.M. Calabrese, E. Hernández-García, C.E. Tarnita, C. López, and J.A. Bonachela. Integrating theory and experiments to link local mechanisms and ecosystem-level consequences of vegetation patterns in drylands.Chaos, Solitons & Fractals, 166:112881, 2023
work page 2023
- [35]
-
[36]
S. Mazzoleni, G. Bonanomi, F. Giannino, G. Incerti, S.C. Dekker, and M. Rietkerk. Modelling the effects of litter decomposition on tree diversity patterns.Ecological Modelling, 221(23):2784–2792, 2010
work page 2010
-
[37]
S. Mazzoleni, G. Bonanomi, G. Incerti, M.L. Chiusano, V. Lanzotti, et al. Inhibitory and toxic effects of extracellular self-DNA in litter: a mechanism for negative plant-soil feedbacks?New Phytologist, 205(3):1195–1210, 2014
work page 2014
-
[38]
E. Meron. Pattern-formation approach to modelling spatially extended ecosystems. Ecological Modelling, 234:70–82, 2012
work page 2012
- [39]
-
[40]
M.Rietkerk, R.Bastiaansen, S.Banerjee, J.vandeKoppel, M.Baudena, andA.Doelman. Evasion of tipping in complex systems through spatial pattern formation.Science, 374(6564):eabj0359, 2021
work page 2021
-
[41]
J.S. Ritchie, A.L. Krause, and R.A. Van Gorder. Turing and wave instabilities in hyperbolic reaction–diffusion systems: The role of second-order time derivatives and cross-diffusion terms on pattern formation. Annals of Physics, 444:169033, 2022
work page 2022
-
[42]
V. Rottschäfer and A. Doelman. On the transition from the Ginzburg-Landau equation to the extended Fisher-Kolmogorov equation.Physica D: Nonlinear Phenomena, 118(3-4):261–292, 1998. 16
work page 1998
-
[43]
N. Salvatori, M. Moreno, M. Zotti, A. Iuorio, F. Cartenì, G. Bonanomi, S. Mazzoleni, and F. Gian- nino. Process based modelling of plants-fungus interactions explains fairy ring types and dynamics. Scientific Reports, 13(1):19918, 2023
work page 2023
- [44]
-
[45]
N. Shigesada, K. Kawasaki, and E. Teramoto. Spatial segregation of interacting species.Journal of Theoretical Biology, 79(1):83–99, 1979
work page 1979
- [46]
-
[47]
H. Uecker. Hopf bifurcation and time periodic orbits withpde2path– Algorithms and applications. Communications in Computational Physics, 25(3):812–852, 2019
work page 2019
-
[48]
H. Uecker. Numerical Continuation and Bifurcation in Nonlinear PDEs. Society for Industrial and Applied Mathematics, 2021
work page 2021
-
[49]
E. Villar-Sepúlveda, A.R. Champneys, and A.L. Krause. Designing reaction-cross-diffusion sys- tems with Turing and wave instabilities.arXiv preprint arXiv:2409.06860, 2024
-
[50]
J. von Hardenberg, E. Meron, M. Shachak, and Y. Zarmi. Diversity of vegetation patterns and desertification. Physical Review Letters, 87(19):198101, 2001
work page 2001
-
[51]
C. Yang, J. Li, F. Zhang, N. Liu, and Y. Zhang. The optimal Redfield N: P ratio caused by fairy ring fungi stimulates plant productivity in the temperate steppe of China.Fungal Ecology, 34:91–98, 2018
work page 2018
- [52]
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.