Graphon Signal Processing for Spiking and Biological Neural Networks
Pith reviewed 2026-05-18 21:50 UTC · model grok-4.3
The pith
Graphon signal processing produces trial-invariant low-dimensional embeddings from neural activity that classify stimuli more accurately than PCA and discrete graph methods.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
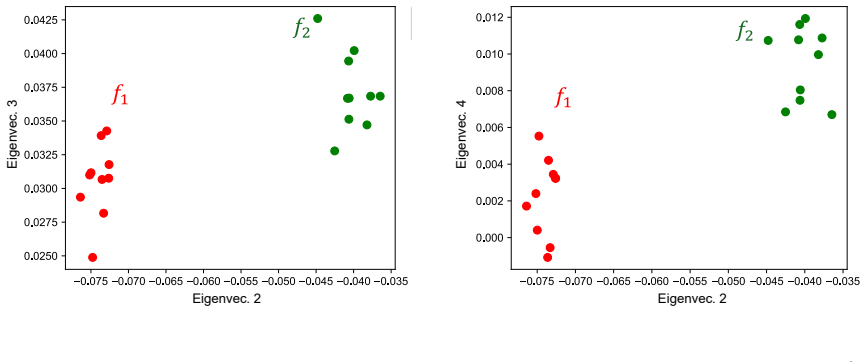
Graphon-based spectral projections yield trial-invariant, low-dimensional embeddings that improve stimulus classification over Principal Component Analysis and discrete GSP baselines. The embeddings remain stable under variations in network stochasticity, providing robustness to different network sizes and noise levels. This framework is applied first to simulated spiking networks and then to calcium imaging recordings from biological networks.
What carries the argument
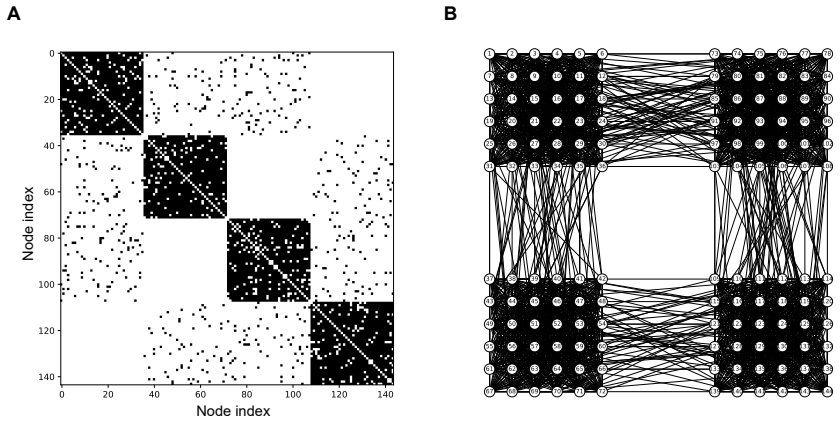
The graphon, a measurable function on the unit square that represents the limit of convergent graph sequences, supplies the spectral decomposition used to project network outputs into stable, low-dimensional coordinates for the inverse problem.
If this is right
- The embeddings remain stable when network stochasticity, size, or noise level changes.
- Stimulus classification accuracy exceeds that of principal component analysis and discrete graph signal processing in both simulated and recorded data.
- The method extends graph signal processing to limits of large networks without requiring explicit construction of the full adjacency matrix.
- This constitutes the first reported use of graphon signal processing on biological neural network data.
Where Pith is reading between the lines
- The same graphon projections could be tested on other inverse problems such as inferring connectivity from observed dynamics in non-neural biological networks.
- Because the embeddings are low-dimensional and stable, they might serve as features for real-time decoding in brain-machine interfaces where computational cost must remain low.
- If the graphon model holds, the approach predicts that classification performance will improve further as network recordings grow larger, unlike methods that degrade with scale.
Load-bearing premise
The connectivity and dynamics of the spiking and biological neural networks can be accurately captured by a graphon model whose spectral properties directly support stable, low-dimensional projections for the stimulus identification inverse problem.
What would settle it
A collection of biological neural recordings in which graphon-derived embeddings produce no gain in stimulus classification accuracy over PCA and lose stability when network noise or size is increased would falsify the central claim.
Figures








read the original abstract
Graph Signal Processing (GSP) extends classical signal processing to signals defined on graphs, enabling filtering, spectral analysis, and sampling of data generated by networks of various kinds. Graphon Signal Processing (GnSP) develops this framework further by employing the theory of graphons. Graphons are measurable functions on the unit square that represent graphs and limits of convergent graph sequences. The use of graphons provides stability of GSP methods to stochastic variability in network data and improves computational efficiency for very large networks. We use GnSP to address the stimulus identification problem (SIP) in computational and biological neural networks. The SIP is an inverse problem that aims to infer the unknown stimulus s from the observed network output f. We first validate the approach in spiking neural network simulations and then analyze calcium imaging recordings. Graphon-based spectral projections yield trial-invariant, lowdimensional embeddings that improve stimulus classification over Principal Component Analysis and discrete GSP baselines. The embeddings remain stable under variations in network stochasticity, providing robustness to different network sizes and noise levels. To the best of our knowledge, this is the first application of GnSP to biological neural networks, opening new avenues for graphon-based analysis in neuroscience.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops Graphon Signal Processing (GnSP) to solve the stimulus identification problem (SIP) in spiking neural network simulations and calcium imaging recordings from biological networks. It claims that spectral projections derived from a graphon yield trial-invariant low-dimensional embeddings that improve stimulus classification accuracy relative to PCA and discrete GSP baselines while remaining stable under changes in network size, stochastic realizations, and noise levels. The work positions itself as the first application of GnSP to biological neural data.
Significance. If the central claims are substantiated with quantitative evidence, the paper would be significant for extending GSP theory to graphon limits in a new domain, offering a route to robust, scalable analysis of large neural networks. The emphasis on stability to stochastic variability and computational efficiency for inverse problems like SIP could influence future work on connectivity inference and stimulus decoding in neuroscience.
major comments (2)
- [§3.1] §3.1 (graphon model definition): the claim that a single graphon spectral decomposition yields trial-invariant embeddings for the SIP rests on the unproven assumption that finite spiking networks converge to the graphon in a cut-norm sense that preserves the low-frequency eigenmodes acting on localized stimulus-driven signals; directed or modular connectivity typical of biological networks may violate this, and no numerical check of eigenmode stability across realizations is provided.
- [Results] Results section on calcium imaging and spiking simulations: the reported improvements in classification and stability lack quantitative metrics (accuracy values, standard deviations, or statistical comparisons) and exclusion criteria, making it impossible to evaluate whether the embeddings genuinely outperform discrete GSP or remain robust when network inhomogeneities are present.
minor comments (2)
- [Abstract] Abstract: the phrase 'improved stimulus classification' should be accompanied by the specific performance metric and effect size even in the abstract.
- [Notation] Notation: the mapping from the observed network output f to the graphon signal is introduced without an explicit equation linking the spiking dynamics to the graphon integral operator.
Simulated Author's Rebuttal
We thank the referee for their constructive and detailed comments, which highlight important aspects for strengthening the theoretical and empirical foundations of our work. We address each major comment point by point below, indicating the revisions we will incorporate.
read point-by-point responses
-
Referee: [§3.1] §3.1 (graphon model definition): the claim that a single graphon spectral decomposition yields trial-invariant embeddings for the SIP rests on the unproven assumption that finite spiking networks converge to the graphon in a cut-norm sense that preserves the low-frequency eigenmodes acting on localized stimulus-driven signals; directed or modular connectivity typical of biological networks may violate this, and no numerical check of eigenmode stability across realizations is provided.
Authors: We acknowledge that a complete theoretical proof of cut-norm convergence preserving the relevant eigenmodes for directed or modular networks is not provided and lies beyond the scope of the current manuscript. The graphon framework is applied here via an undirected approximation derived from absolute connectivity strengths, which is standard in initial GnSP applications to neural data. To directly address the lack of numerical verification, the revised manuscript will include new experiments showing the stability of the low-frequency eigenmodes (e.g., eigenvalue spectra and eigenvector alignments) across multiple stochastic realizations, network sizes, and stimulus conditions. These checks will quantify variation using metrics such as subspace angles and will be presented in an expanded Section 3 or supplementary material. revision: partial
-
Referee: [Results] Results section on calcium imaging and spiking simulations: the reported improvements in classification and stability lack quantitative metrics (accuracy values, standard deviations, or statistical comparisons) and exclusion criteria, making it impossible to evaluate whether the embeddings genuinely outperform discrete GSP or remain robust when network inhomogeneities are present.
Authors: We agree that the results would be more convincing with explicit quantitative details. In the revised manuscript, we will expand the results section to report mean classification accuracies with standard deviations across trials for GnSP, PCA, and discrete GSP baselines. Statistical comparisons (e.g., paired t-tests with p-values) will be added to quantify improvements. For the calcium imaging experiments, we will explicitly describe the exclusion criteria applied to recordings and neurons (e.g., signal-to-noise thresholds and trial selection rules). These additions will also include robustness checks under controlled inhomogeneities to better demonstrate stability. revision: yes
Circularity Check
No circularity: derivation applies external graphon theory to new domain
full rationale
The paper's central claims rest on applying established graphon signal processing (GnSP) theory—developed independently in prior literature—to spiking and biological neural networks for the stimulus identification problem. The abstract and described method use graphon spectral projections to obtain embeddings, then compare them empirically to PCA and discrete GSP baselines on simulation and calcium imaging data. No equations or steps are shown to define a quantity in terms of itself, fit a parameter on a subset and relabel the output as a prediction, or rely on a self-citation chain whose validity is presupposed by the present work. The reported improvements and stability are presented as empirical outcomes rather than algebraic identities forced by construction. The derivation chain therefore remains self-contained against external benchmarks and does not reduce to its inputs.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Spiking and biological neural networks can be modeled as convergent graph sequences whose limit is a graphon whose spectral properties yield stable low-dimensional embeddings for stimulus identification.
Lean theorems connected to this paper
-
IndisputableMonolith/Foundation/AbsoluteFloorClosure.leanreality_from_one_distinction unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
Graphon-based spectral projections yield trial-invariant, lowdimensional embeddings that improve stimulus classification over Principal Component Analysis and discrete GSP baselines. The embeddings remain stable under variations in network stochasticity
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
We define W(x,y)=∑Cij 1Ii(x)1Ij(y) and compute eigenvalues λ1=1/4, λ2=λ3=(1−2α)/4 together with piecewise-constant eigenfunctions ϕi
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
G. Leus, A. G. Marques, J. M. F. Moura, A. Ortega, and D. I. Shuman. Graph signal processing: History, development, impact, and outlook. IEEE Signal Processing Magazine, 40(4):49–60, 2023
work page 2023
- [2]
-
[3]
D. I. Shuman, S. K. Narang, P. Frossard, A. Ortega, and P. Vandergheynst. The emerging field of signal processing on graphs: Extending high-dimensional data analysis to networks and other irregular domains. IEEE Signal Processing Magazine, 30(3):83–98, 2013
work page 2013
-
[4]
L. Ruiz, L. F. O. Chamon, and A. Ribeiro. Graphon signal processing. IEEE Transactions on Signal Processing, 69:4961–4976, 2021. 17
work page 2021
- [5]
-
[6]
M. Montal `a-Flaquer, C. F. L´opez-Le´on, D. Tornero, A. M. Houben, T. Fardet, P. Monceau, S. Bottani, and J. Soriano. Rich dynamics and functional organization on topographically designed neuronal networks in vitro. iScience, 25(12):105680, 2022
work page 2022
-
[7]
T. Sumi, A. M. Houben, H. Yamamoto, H. Kato, Y . Katori, J. Soriano, and A. Hirano-Iwata. Modular architecture confers robustness to damage and facilitates recovery in spiking neural networks modeling in vitro neurons. Frontiers in Neuroscience, 19:1570783, 2025
work page 2025
-
[8]
A. K. V ogt, L. Lauer, W. Knoll, and A. Offenh ¨ausser. Micropatterned substrates for the growth of functional neuronal networks of defined geometry. Biotechnology Progress, 19(5):1562–1568, 2003
work page 2003
-
[9]
J. Albers and A. Offenh ¨ausser. Signal propagation between neuronal populations controlled by mi- cropatterning. Frontiers in Bioengineering and Biotechnology, 4, 2016
work page 2016
-
[10]
H. Yamamoto, R. Matsumura, H. Takaoki, S. Katsurabayashi, A. Hirano-Iwata, and M. Niwano. Uni- directional signal propagation in primary neurons micropatterned at a single-cell resolution. Applied Physics Letters, 109(4):043703, 2016
work page 2016
-
[11]
H. Yamamoto, S. Moriya, K. Ide, T. Hayakawa, H. Akima, S. Sato, S. Kubota, T. Tanii, M. Niwano, S. Teller, J. Soriano, and A. Hirano-Iwata. Impact of modular organization on dynamical richness in cortical networks. Science Advances, 4(11):eaau4914, 2018
work page 2018
-
[12]
L. J. Millet and M. U. Gillette. New perspectives on neuronal development via microfluidic environ- ments. Trends in Neurosciences, 35(12):752–761, 2012. Epub 2012 Sep 29
work page 2012
- [13]
-
[14]
L. Pan, S. Alagapan, E. Franca, S. S. Leondopulos, T. B. DeMarse, G. J. Brewer, and B. C. Wheeler. An in vitro method to manipulate the direction and functional strength between neural populations. Frontiers in Neural Circuits, 9, 2015
work page 2015
-
[15]
R. Habibey, J. Striebel, M. Meinert, R. Latiftikhereshki, F. Schmieder, R. Nasiri, and S. Latifi. Engi- neered modular neuronal networks-on-chip represent structure-function relationship. Biosensors and Bioelectronics, 261:116518, 2024
work page 2024
-
[16]
N. Winter-Hjelm, ˚A. Brune Tomren, P. Sikorski, A. Sandvig, and I. Sandvig. Structure-function dynamics of engineered, modular neuronal networks with controllable afferent-efferent connectivity. Journal of Neural Engineering, 20(4):046024, 2023
work page 2023
-
[17]
T. Sumi, H. Yamamoto, Y . Katori, K. Ito, S. Moriya, T. Konno, S. Sato, and A. Hirano-Iwata. Biolog- ical neurons act as generalization filters in reservoir computing. Proceedings of the National Academy of Sciences, 120(27):e2217008120, 2023
work page 2023
-
[18]
D. Meunier, R. Lambiotte, and E. T. Bullmore. Modular and hierarchically modular organization of brain networks. Frontiers in Neuroscience, 4, 2010. 18
work page 2010
-
[19]
O. Sporns and R. F. Betzel. Modular brain networks. Annual Review of Psychology, 67(1):613–640, 2016
work page 2016
-
[20]
N. Biggs. Algebraic graph theory. Camb. Math. Libr. Cambridge: Cambridge University Press, 2nd ed. edition, 1994
work page 1994
-
[21]
A. Ortega. Introduction to graph signal processing. Cambridge: Cambridge University Press, 2022
work page 2022
-
[22]
M. W. Morency and G. Leus. Graphon filters: Graph signal processing in the limit. IEEE Transactions on Signal Processing, 69:1740–1754, 2021
work page 2021
-
[23]
N. Young. An introduction to Hilbert space . Camb. Math. Textb. Cambridge (UK) etc.: Cambridge University Press, 1988
work page 1988
-
[24]
M. Ghandehari and G. S. Medvedev. The Large Deviation Principle for W -random spectral measures. Appl. Comput. Harmon. Anal., 77:12, 2025. Id/No 101756
work page 2025
-
[25]
D. A. French and E. I. Gruenstein. An integrate-and-fire model for synchronized bursting in a network of cultured cortical neurons. Journal of Computational Neuroscience, 21(3):227–241, 2006
work page 2006
-
[26]
Y . Ishikawa, T. Shinkawa, T. Sumi, H. Kato, H. Yamamoto, and Y . Katori. Integrating predictive coding with reservoir computing in spiking neural network model of cultured neurons. Nonlinear Theory and Its Applications, IEICE, 15(2):432–442, 2024
work page 2024
-
[27]
Y . Sato, H. Yamamoto, Y . Ishikawa, T. Sumi, Y . Sono, S. Sato, Y . Katori, and A. Hirano-Iwata. In silico modeling of reservoir-based predictive coding in biological neuronal networks on microelectrode arrays. Japanese Journal of Applied Physics, 63(10):108001, 2024
work page 2024
-
[28]
T. Sumi, H. Yamamoto, Y . Katori, K. Ito, S. Moriya, T. Konno, S. Sato, and A. Hirano-Iwata. Dataset for: Biological neurons act as generalization filters in reservoir computing. Zenodo, 2023. Dataset
work page 2023
-
[29]
J. P. Cunningham and B. M. Yu. Dimensionality reduction for large-scale neural recordings. Nature Neuroscience, 17:1500–1509, 2014
work page 2014
- [30]
- [31]
- [32]
-
[33]
P. J. Thomas, A. Leow, H. Klumpp, K. L. Phan, and O. Ajilore. Default mode network hypoalignment of function to structure correlates with depression and rumination. Biological Psychiatry: Cognitive Neuroscience and Neuroimaging, 9(1):101–111, 2024. 19
work page 2024
-
[34]
I. Rigoni, J. Ru ´e-Queralt, K. Glomb, M. G. Preti, N. Roehri, S. Tourbier, L. Spinelli, S. Vulliemoz, G. Plomp, and D. Van De Ville. Structure–function coupling increases during interictal spikes in temporal lobe epilepsy: A graph signal processing study. Clinical Neurophysiology, 153:1–10, 2023
work page 2023
-
[35]
J. I. Glaser, A. S. Benjamin, R. H. Chowdhury, M. G. Perich, L. E. Miller, and K. P. Kording. Machine learning for neural decoding. eNeuro, 7(4):ENEURO.0506–19.2020, 2020
work page 2020
- [36]
- [37]
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.