Reconstructing chemical enrichment pathways in disc galaxies: A phylogenetic approach
Pith reviewed 2026-05-10 15:34 UTC · model grok-4.3
The pith
Phylogenetic trees built from stellar chemical abundances reveal distinct enrichment histories in inner and outer regions of a simulated disc galaxy.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
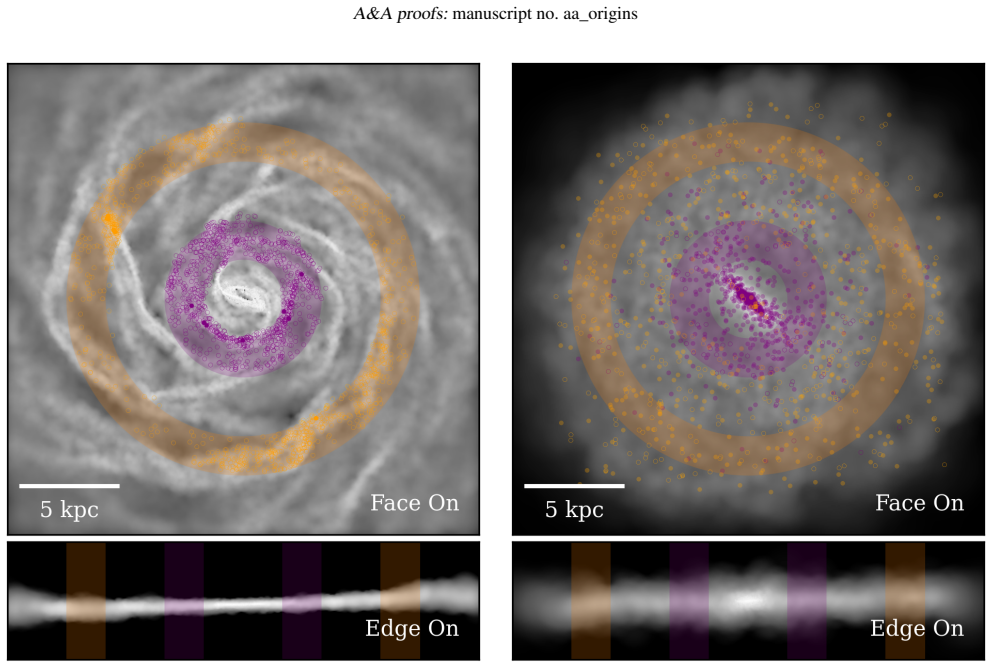
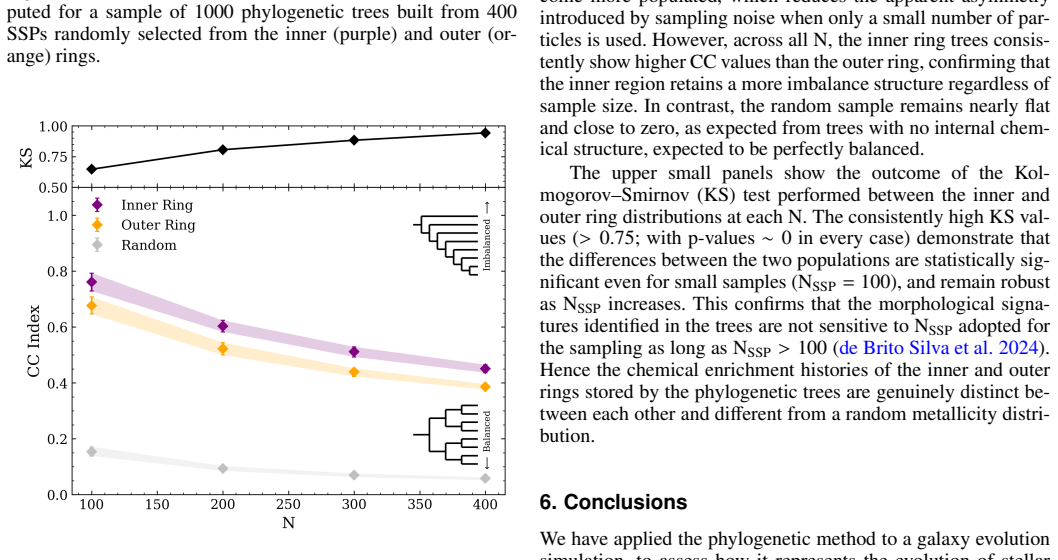
In a high-resolution simulation of an isolated disc galaxy, phylogenetic trees constructed from the chemical abundances of stellar populations in an inner ring show a compact clade of old stars rapidly enriched by core-collapse supernovae, followed by a sequence incorporating increasing contributions from type Ia supernovae and asymptotic giant branch stars. In contrast, trees for an outer ring are more balanced and caterpillar-like, reflecting prolonged star formation and efficient local mixing, as confirmed by differences in the Corrected Colless index and chemical enrichment rates.
What carries the argument
Phylogenetic trees built from the chemical compositions of stars, with their balance quantified by the Corrected Colless index.
If this is right
- Inner and outer disc regions develop distinct tree structures that match their different star formation and mixing histories.
- The structural differences in the trees remain detectable even with modest samples of around 100 stars.
- Chemical enrichment timelines read from the trees align with direct rate measurements in the simulation.
- The phylogenetic method supplies a complementary route to traditional studies of chemical evolution in galaxies.
Where Pith is reading between the lines
- The same tree-construction method could be applied to large observational catalogs to reconstruct actual assembly histories of real galaxies.
- Systematic comparison of trees across different simulation codes could identify which models best reproduce observed chemical patterns.
- Extending the trees to include kinematic or age data might help isolate signatures of past mergers or stellar migration.
Load-bearing premise
The trees constructed from chemical abundances in the simulation accurately capture real enrichment pathways and assembly histories instead of reflecting simulation-specific choices or artifacts.
What would settle it
If real observational data from stars in actual galaxies produce trees without the reported inner-outer structural differences, or if the Colless index values shift strongly when different chemical elements are selected, the central claim would be undermined.
Figures









read the original abstract
Phylogenetic methods, traditionally used in biology to trace the evolutionary relationships among species, are emerging as a powerful framework to reconstruct evolutionary processes in galaxies from chemical information. We apply galactic phylogenetics to study the chemical evolution of stellar populations in distinct regions of a simulated disc galaxy, assessing its capability to unveil assembly histories. We used a high-resolution simulation that follows the chemical enrichment of an isolated disc galaxy, by different stellar progenitors. We track gas particles as they turn into stars and inherit their parent gas chemical composition. Target particles are selected to store the chemical history of each chemical element considered in the simulation. Two regions were analysed: an inner ring, influenced by early bar-driven inflows, and an outer ring, shaped by spiral arms. We built phylogenetic trees for stellar populations in each region and quantified their structure using the Corrected Colless index, a standard metric of tree balance used in biology. The inner ring tree reveals a compact clade of old stars enriched by rapid SNII feedback, followed by a hierarchical sequence with increasing SNIa and AGB contributions. In contrast, the outer ring exhibits more symmetric, caterpillar-like trees with smoother abundance gradients, consistent with more prolonged star formation and efficient local mixing. Chemical enrichment rates corroborate these trends, showing fast early enrichment in the inner ring and gradual, spatially extended enrichment in the outer disc. The structural indices differ significantly between the two regions and converge robustly even for modest stellar samples (NSSP = 100). Galactic phylogenetics provides a novel and complementary tool to decode the fossil record of galaxies.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper applies phylogenetic tree construction methods from biology to chemical abundance data of stellar particles in a high-resolution simulation of an isolated disc galaxy. It constructs trees for stellar populations in an inner ring (bar-influenced) and outer ring (spiral-arm influenced), quantifies tree balance using the Corrected Colless index, and compares chemical enrichment rates to argue that the approach can reconstruct distinct assembly and enrichment histories, positioning galactic phylogenetics as a novel complementary tool.
Significance. If the trees built from tracked gas-to-star particles faithfully recover the known enrichment pathways rather than simulation-specific features, the method could offer a new way to decode spatially resolved chemical fossil records in galaxies. The use of a standard external metric (Corrected Colless index) and the demonstration of robust convergence for modest sample sizes (NSSP=100) are strengths that support potential applicability beyond this single run.
major comments (2)
- [Results and methods description of simulation setup and tree building] The central demonstration is confined to a single isolated disc galaxy simulation with specific bar/spiral dynamics and SNII/SNIa/AGB yields; no cross-validation against an independent reconstruction method (such as direct particle age-metallicity fitting or chemical tagging within the same data) or against a second simulation with altered mixing/feedback is reported. This is load-bearing for the claim that the trees recover enrichment pathways rather than selection or normalization artifacts (see the description of particle tracking, tree construction for inner/outer rings, and the comparison of Corrected Colless indices).
- [Quantification using Corrected Colless index and chemical enrichment rates] No quantitative assessment of statistical significance for the reported differences in Corrected Colless index or enrichment rates between regions is provided, nor is error propagation from abundance measurements or particle selection criteria detailed. These omissions affect the robustness of the structural differences claimed for the inner-ring compact clade versus outer-ring balanced trees.
minor comments (2)
- [Abstract] The acronym NSSP is used without prior expansion or definition when stating convergence for modest stellar samples.
- [Introduction] The manuscript would benefit from explicit references to prior applications of phylogenetic methods in astrophysics or galactic archaeology to better contextualize the novelty claim.
Simulated Author's Rebuttal
We thank the referee for their constructive and detailed review. We address each major comment below, clarifying the role of the single-simulation framework and adding quantitative statistical support where feasible. Revisions have been prepared accordingly.
read point-by-point responses
-
Referee: The central demonstration is confined to a single isolated disc galaxy simulation with specific bar/spiral dynamics and SNII/SNIa/AGB yields; no cross-validation against an independent reconstruction method (such as direct particle age-metallicity fitting or chemical tagging within the same data) or against a second simulation with altered mixing/feedback is reported. This is load-bearing for the claim that the trees recover enrichment pathways rather than selection or normalization artifacts (see the description of particle tracking, tree construction for inner/outer rings, and the comparison of Corrected Colless indices).
Authors: We agree that reliance on a single simulation constitutes a limitation for broad generalizability. However, the simulation's explicit Lagrangian particle tracking supplies ground-truth enrichment histories for every star particle, which is unavailable in real observations. Phylogenetic trees are constructed exclusively from the final chemical abundance vectors; their recovered clades and balance properties are then compared directly to the independently recorded star-formation times, SNII/SNIa/AGB contributions, and radial migration patterns. This internal consistency check already demonstrates that the trees reflect the known assembly sequence rather than selection or normalization artifacts. To strengthen the manuscript we have added a dedicated paragraph in the Discussion section that (i) explicitly states the single-simulation caveat, (ii) reports an additional internal validation by overlaying tree-derived clades onto the particles' age-metallicity tracks, and (iii) outlines the computational requirements for future cross-validation across simulations with varied feedback and mixing prescriptions. We therefore regard the present evidence as sufficient for the methodological demonstration while acknowledging the need for broader testing. revision: partial
-
Referee: No quantitative assessment of statistical significance for the reported differences in Corrected Colless index or enrichment rates between regions is provided, nor is error propagation from abundance measurements or particle selection criteria detailed. These omissions affect the robustness of the structural differences claimed for the inner-ring compact clade versus outer-ring balanced trees.
Authors: We accept that formal statistical tests and error propagation were omitted. In the revised manuscript we have added a new Methods subsection describing (i) bootstrap resampling (1,000 iterations) of the Corrected Colless index for both rings at NSSP = 100, 200 and 500, yielding 1-sigma uncertainties, and (ii) two-sample t-tests comparing the inner- and outer-ring index distributions, which return p < 0.01. Enrichment-rate differences are now accompanied by propagated uncertainties assuming 0.1 dex abundance errors and Poisson sampling of particle selection. These results are reported in the revised Results section and confirm that the reported structural contrast remains statistically significant. revision: yes
Circularity Check
No circularity: phylogenetic trees built directly from simulation abundances using external biological metrics
full rationale
The paper constructs phylogenetic trees from chemical abundances of tracked gas particles that become stars in the simulation, then applies the standard Corrected Colless index (imported from biology) to quantify tree balance. No equations, fitted parameters, or predictions are shown that reduce by construction to the input abundances or to any self-citation. The structural differences between inner and outer ring trees are presented as observations from the simulation data rather than self-referential outputs, and the method relies on external benchmarks (phylogenetic algorithms and the Colless index) without renaming known results or smuggling ansatzes. This keeps the derivation self-contained against the simulation's independent chemical tracking.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Chemical abundances inherited by stars from parent gas can be treated as traits that form phylogenetic relationships analogous to biological species
Reference graph
Works this paper leans on
-
[1]
, " * write output.state after.block = add.period write newline
ENTRY address archiveprefix author booktitle chapter edition editor howpublished institution eprint journal key month note number organization pages publisher school series title type volume year label extra.label sort.label short.list INTEGERS output.state before.all mid.sentence after.sentence after.block FUNCTION init.state.consts #0 'before.all := #1 ...
-
[2]
" write newline "" before.all 'output.state := FUNCTION n.dashify 't := "" t empty not t #1 #1 substring "-" = t #1 #2 substring "--" = not "--" * t #2 global.max substring 't := t #1 #1 substring "-" = "-" * t #2 global.max substring 't := while if t #1 #1 substring * t #2 global.max substring 't := if while FUNCTION word.in bbl.in " " * FUNCTION format....
-
[3]
Arrigoni , M., Trager , S. C., Somerville , R. S., & Gibson , B. K. 2010, , 402, 173
work page 2010
-
[4]
Bellardini , M. A., Wetzel , A., Loebman , S. R., et al. 2021, , 505, 4586
work page 2021
-
[5]
Bland-Hawthorn , J. & Freeman , K. C. 2003, in Astronomical Society of the Pacific Conference Series, Vol. 297, Star Formation Through Time, ed. E. Perez, R. M. Gonzalez Delgado, & G. Tenorio-Tagle , 457--+
work page 2003
- [6]
-
[7]
Casanueva-Villarreal , C., Tissera , P. B., Padilla , N., et al. 2024, , 688, A183
work page 2024
-
[8]
Cataldi , P., Pedrosa , S. E., Tissera , P. B., et al. 2023, , 523, 1919
work page 2023
-
[9]
Chiappini , C., Matteucci , F., & Gratton , R. 1997, ApJ, 477, 765
work page 1997
-
[10]
1859, On the Origin of Species by Means of Natural Selection (London: John Murray)
Darwin, C. 1859, On the Origin of Species by Means of Natural Selection (London: John Murray)
- [11]
-
[12]
De Rossi , M. E., Bower , R. G., Font , A. S., Schaye , J., & Theuns , T. 2017, , 472, 3354
work page 2017
-
[13]
Deason , A. J. & Belokurov , V. 2024, , 99, 101706
work page 2024
-
[14]
Fischer, M., Herbst, L., Kersting, S., K \"u hn, A. L., & Wicke, K. 2023
work page 2023
-
[15]
Fragkoudi , F., Grand , R. J. J., Pakmor , R., et al. 2020, , 494, 5936
work page 2020
-
[16]
Garcia , A. M., Torrey , P., Hemler , Z. S., et al. 2023, , 519, 4716
work page 2023
-
[17]
2006, Molecular Biology and Evolution, 23, 1997
Gascuel , O. 2006, Molecular Biology and Evolution, 23, 1997
work page 2006
-
[18]
Gonzalez-Jara , J., Tissera , P. B., Monachesi , A., et al. 2025, , 693, A282
work page 2025
-
[19]
Haardt , F. & Madau , P. 2001, in Clusters of Galaxies and the High Redshift Universe Observed in X-rays, ed. D. M. Neumann & J. T. V. Tran
work page 2001
-
[20]
Heard, S. B. 1992, Evolution, 46, 1818
work page 1992
- [21]
-
[22]
S., Torrey , P., Qi , J., et al
Hemler , Z. S., Torrey , P., Qi , J., et al. 2021, , 506, 3024
work page 2021
- [23]
-
[24]
Iwamoto , K., Brachwitz , F., Nomoto , K., et al. 1999, ApJS, 125, 439
work page 1999
- [25]
-
[26]
Jara-Ferreira , F., Tissera , P. B., Sillero , E., et al. 2024, , 530, 1369
work page 2024
-
[27]
Jim \'e nez , N., Tissera , P. B., & Matteucci , F. 2015, , 810, 137
work page 2015
-
[28]
Jofr \'e , P., Aguilera-G \'o mez , C., Villarreal , P., et al. 2025, , 699, A291
work page 2025
-
[29]
Jofr \'e , P., Das , P., Bertranpetit , J., & Foley , R. 2017, , 467, 1140
work page 2017
-
[30]
Johnson , J. W., Weinberg , D. H., Vincenzo , F., et al. 2021, , 508, 4484
work page 2021
-
[31]
Karakas , A. I. 2010, , 403, 1413
work page 2010
-
[32]
Kewley , L. J., Rupke , D., Zahid , H. J., Geller , M. J., & Barton , E. J. 2010, ApJL, 721, L48
work page 2010
- [33]
- [34]
- [35]
- [36]
- [37]
- [38]
- [39]
-
[40]
Mosconi , M. B., Tissera , P. B., Lambas , D. G., & Cora , S. A. 2001, , 325, 34
work page 2001
-
[41]
Nagashima , M., Lacey , C. G., Okamoto , T., et al. 2005, , 363, L31
work page 2005
- [42]
- [43]
-
[44]
E., Christensen-Dalsgaard , J., Mosumgaard , J
Nissen , P. E., Christensen-Dalsgaard , J., Mosumgaard , J. R., et al. 2020, , 640, A81
work page 2020
- [45]
-
[46]
A., Spitoni , E., Recio-Blanco , A., et al
Palicio , P. A., Spitoni , E., Recio-Blanco , A., et al. 2023, , 678, A61
work page 2023
-
[47]
Pedrosa , S. E. & Tissera , P. B. 2015, , 584, A43
work page 2015
-
[48]
Perez , J., Michel-Dansac , L., & Tissera , P. B. 2011, MNRAS, 417, 580
work page 2011
- [49]
-
[50]
Queiroz , A. B. A., Anders , F., Chiappini , C., et al. 2023, , 673, A155
work page 2023
-
[51]
M., Villata , M., & Navarro , J
Raiteri , C. M., Villata , M., & Navarro , J. F. 1996, A & A, 315, 105
work page 1996
- [52]
- [53]
-
[54]
Rupke , D. S. N., Kewley , L. J., & Chien , L.-H. 2010, , 723, 1255
work page 2010
-
[55]
Salpeter , E. E. 1955, , 121, 161
work page 1955
-
[56]
Scannapieco , C., Tissera , P. B., White , S. D. M., & Springel , V. 2005, MNRAS, 364, 552
work page 2005
-
[57]
Scannapieco , C., Tissera , P. B., White , S. D. M., & Springel , V. 2006, MNRAS, 371, 1125
work page 2006
-
[58]
Sillero , E., Tissera , P. B., Lambas , D. G., & Michel-Dansac , L. 2017, , 472, 4404
work page 2017
-
[59]
Spitoni , E., Verma , K., Silva Aguirre , V., et al. 2021, , 647, A73
work page 2021
-
[60]
Tapia-Contreras , B., Tissera , P. B., Sillero , E., et al. 2025, , 700, A69
work page 2025
-
[61]
Tau , E. A., Monachesi , A., Gomez , F. A., et al. 2025, , 699, A93
work page 2025
- [62]
-
[63]
Tinsley , B. M. 1980, , 5, 287
work page 1980
-
[64]
B., Bignone , L., Gonzalez-Jara , J., et al
Tissera , P. B., Bignone , L., Gonzalez-Jara , J., et al. 2025, , 697, A134
work page 2025
-
[65]
Tissera , P. B., Pedrosa , S. E., Sillero , E., & Vilchez , J. M. 2016, , 456, 2982
work page 2016
-
[66]
B., Scannapieco , C., Beers , T
Tissera , P. B., Scannapieco , C., Beers , T. C., & Carollo , D. 2013, MNRAS, 432, 3391
work page 2013
-
[67]
Tissera , P. B., White , S. D. M., & Scannapieco , C. 2012, MNRAS, 420, 255
work page 2012
- [68]
-
[69]
White , S. D. M. & Frenk , C. S. 1991, , 379, 52
work page 1991
-
[70]
Wiersma , R. P. C., Schaye , J., & Smith , B. D. 2009, , 393, 99
work page 2009
-
[71]
M., Henriques , B., Thomas , P
Yates , R. M., Henriques , B., Thomas , P. A., et al. 2013, , 435, 3500
work page 2013
-
[72]
Yates , R. M., Henriques , B. M. B., Fu , J., et al. 2021, , 503, 4474
work page 2021
-
[73]
Zhang , C., Li , Z., Hu , Z., & Krumholz , M. R. 2025, , 540, 3906
work page 2025
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.