Multi-stage volume exclusion models for cell proliferation
Pith reviewed 2026-05-10 01:04 UTC · model grok-4.3
The pith
Multi-stage cell cycle models with myopic proliferation and volume exclusion can be approximated by mean-field PDEs for cell growth and invasion.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
We develop stochastic, on-lattice agent-based models which incorporate volume exclusion, random movement, and multi-stage representations of the cell cycle. The multi-stage framework enables a more realistic representation of true cell cycle time distributions. We also introduce a novel form of myopic behaviour, in which cells sense their local environment when attempting to proliferate. For each ABM, we derive a corresponding continuum partial differential equation description under the mean-field approximation. Using numerical simulations, we investigate how different proliferation mechanisms influence population-level dynamics in both the discrete and continuum models in growth-to-conflue
What carries the argument
The multi-stage cell cycle framework in volume-excluding on-lattice agent-based models with myopic proliferation sensing.
Load-bearing premise
The mean-field approximation holds so that correlations between nearby cells do not significantly affect the average population behavior.
What would settle it
A large mismatch between the solutions of the derived PDEs and the averaged outcomes from repeated simulations of the corresponding agent-based models in either confluence growth or invasion wave metrics.
Figures



















read the original abstract
Cell proliferation and cell movement are fundamentally stochastic processes which lead to variability in the growth and spatial structure of cell populations in many biological settings, such as cell invasion, wound healing, and tumour growth. We develop stochastic, on-lattice agent-based models (ABMs) which incorporate volume exclusion, random movement, and multi-stage representations of the cell cycle. The multi-stage framework enables a more realistic representation of true cell cycle time distributions. We also introduce a novel form of myopic behaviour, in which cells sense their local environment when attempting to proliferate. For each ABM, we derive a corresponding continuum partial differential equation (PDE) description under the mean-field approximation. Using numerical simulations, we investigate how different proliferation mechanisms influence population-level dynamics in both the discrete and continuum models. In particular, we consider biologically relevant contexts of growth-to-confluence assays (using uniform initial conditions) and travelling wave behaviour associated with cell invasion. We examine how the PDE solutions compare with the behaviour of the corresponding ABMs averaged over many realisations.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops stochastic on-lattice agent-based models (ABMs) for cell proliferation incorporating volume exclusion, random movement, multi-stage cell-cycle representations (to produce non-exponential division-time distributions), and a novel myopic proliferation rule in which cells sense local occupancy before attempting division. For each ABM variant the authors derive a corresponding continuum PDE under the standard mean-field closure. Numerical simulations then compare ensemble-averaged ABM trajectories against PDE solutions for two biologically relevant settings: growth-to-confluence under uniform initial conditions and travelling-wave invasion.
Significance. If the mean-field PDEs remain quantitatively faithful, the work supplies a practical route to embed realistic cell-cycle timing and local sensing into continuum models of invasion and wound healing, while retaining the computational advantages of PDEs. The explicit ABM-to-PDE derivation and side-by-side numerical tests constitute a clear strength; the multi-stage formulation directly addresses a known limitation of single-stage exponential waiting times.
major comments (2)
- [PDE derivation for myopic proliferation] The mean-field closure used to obtain the PDE proliferation term (standard independence assumption P(occupied_i and occupied_j) = P_i P_j) is applied to the myopic rule, which by construction correlates a cell's division decision with the occupancy of its sensing neighbourhood. Volume exclusion further enforces hard-core correlations. The manuscript does not report any diagnostic (e.g., measured pair-correlation functions from the ABM, comparison of effective proliferation rates, or higher-moment closures) to confirm that the closure remains accurate near fronts or at moderate-to-high densities where these correlations are strongest.
- [Numerical results (growth-to-confluence and travelling waves)] In the numerical comparison sections, ABM results are presented as averages over realisations but without error bars, standard deviations, or convergence tests with respect to lattice size, number of realisations, or sensing radius. Consequently it is impossible to judge whether apparent agreement with the PDE is statistically robust or merely qualitative, especially for the travelling-wave speed and the approach to confluence.
minor comments (2)
- The abstract and introduction would benefit from a concise statement of the precise sensing radius and number of cell-cycle stages used in the main figures, rather than deferring all parameter values to the methods.
- Figure legends should explicitly state the initial density, lattice size, and number of ABM realisations for each panel to allow direct replication of the reported comparisons.
Simulated Author's Rebuttal
We thank the referee for their thoughtful comments on our manuscript. We address each of the major comments below and outline the revisions we plan to make to strengthen the paper.
read point-by-point responses
-
Referee: [PDE derivation for myopic proliferation] The mean-field closure used to obtain the PDE proliferation term (standard independence assumption P(occupied_i and occupied_j) = P_i P_j) is applied to the myopic rule, which by construction correlates a cell's division decision with the occupancy of its sensing neighbourhood. Volume exclusion further enforces hard-core correlations. The manuscript does not report any diagnostic (e.g., measured pair-correlation functions from the ABM, comparison of effective proliferation rates, or higher-moment closures) to confirm that the closure remains accurate near fronts or at moderate-to-high densities where these correlations are strongest.
Authors: We agree that the mean-field approximation may be affected by correlations induced by the myopic proliferation rule and volume exclusion. The manuscript presents the derivation under the standard mean-field closure and shows that the resulting PDEs provide a good approximation to the averaged ABM dynamics in the regimes considered. To directly address this point, we will add diagnostics in the revised manuscript, including computed pair-correlation functions from ABM simulations at various densities and near fronts, to evaluate the validity of the independence assumption. If significant deviations are found, we will discuss their implications for the approximation. revision: yes
-
Referee: [Numerical results (growth-to-confluence and travelling waves)] In the numerical comparison sections, ABM results are presented as averages over realisations but without error bars, standard deviations, or convergence tests with respect to lattice size, number of realisations, or sensing radius. Consequently it is impossible to judge whether apparent agreement with the PDE is statistically robust or merely qualitative, especially for the travelling-wave speed and the approach to confluence.
Authors: We acknowledge that the absence of error bars and convergence tests limits the assessment of the numerical agreement. In the revised version, we will include error bars or standard deviations on the ensemble-averaged ABM results. Additionally, we will perform and report convergence tests with respect to the number of realizations, lattice size, and sensing radius to demonstrate the statistical robustness of the comparisons, particularly for travelling wave speeds and confluence dynamics. revision: yes
Circularity Check
No circularity: standard mean-field derivation from ABMs with independent numerical validation
full rationale
The paper constructs on-lattice ABMs incorporating volume exclusion, random movement, multi-stage cell cycles, and a novel myopic proliferation rule, then applies the standard mean-field closure to obtain continuum PDEs. These PDEs are not obtained by fitting parameters to the same data being predicted, nor by self-definition, renaming, or load-bearing self-citation chains. The subsequent numerical comparisons between averaged ABM realizations and PDE solutions constitute an external check rather than a tautology. No quoted step reduces the claimed result to its inputs by construction.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Mean-field approximation accurately captures the average behaviour of the stochastic ABM
Reference graph
Works this paper leans on
-
[1]
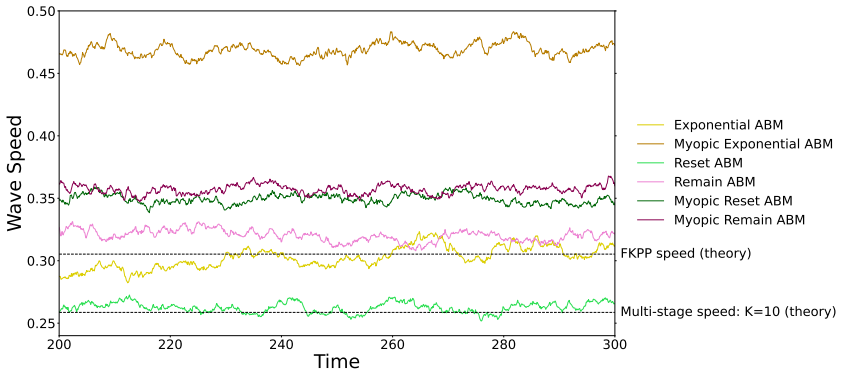
Low motility We begin by considering a regime in which the motility rate,rm, is of the same order as the proliferation rate,rp (previously the low motility regime). In Figure 13, we can see that the wave-fronts which form in each ABM have similar shapes and are very steep. This is because cell proliferation is the major factor driving the travelling wave ...
work page 2000
-
[2]
High motility We now consider a parameter regime in which the motility rate is high relative to the proliferation rate. In Figure 16, we see that even after300time units, the Exponential and Reset models have not yet reached a steady state wavefront. We also note that the wavefronts across all models under this high motility scheme are shallower than in t...
work page 2000
-
[3]
B. Alberts, D. Bray, J. Lewis, M. Raff, K. Roberts, and J.D. Watson.Molecular Biology of the Cell. Garland, 4th edition, 2002
work page 2002
-
[4]
D. Hanahan and R.A. Weinberg. Hallmarks of cancer: the next generation.Cell, 144 (5):646–674, 2011
work page 2011
-
[5]
G.C. Gurtner, S. Werner, Y. Barrandon, and M.T. Longaker. Wound repair and regen- eration.Nature, 453(7193):314–321, 2008
work page 2008
-
[6]
K.A. Schafer. The cell cycle: a review.Veterinary Pathology, 35(6):461–478, 1998
work page 1998
-
[7]
J.A. Pietenpol and Z.A. Stewart. Cell cycle checkpoint signaling: cell cycle arrest versus apoptosis.Toxicology, 181:475–481, 2002. 51
work page 2002
-
[8]
D.G. Johnson and C.L. Walker. Cyclins and cell cycle checkpoints.Annual Review of Pharmacology and Toxicology, 39(1):295–312, 1999
work page 1999
-
[9]
A. Meerson, M. Milyavsky, and V. Rotter. p53 mediates density-dependent growth arrest.FEBS Letters, 559(1-3):152–158, 2004
work page 2004
-
[10]
O.V. Leontieva, Z.N. Demidenko, and M.V. Blagosklonny. Contact inhibition and high cell density deactivate the mammalian target of rapamycin pathway, thus suppressing the senescence program.Proceedings of the National Academy of Sciences, 111(24): 8832–8837, 2014
work page 2014
- [11]
-
[12]
G. An, Q. Mi, J. Dutta-Moscato, and Y. Vodovotz. Agent-based models in translational systems biology.Wiley Interdisciplinary Reviews: Systems Biology and Medicine, 1(2): 159–171, 2009
work page 2009
-
[13]
M. Baar, L. Coquille, H. Mayer, M. Hölzel, M. Rogava, T. Tüting, and A. Bovier. A stochastic model for immunotherapy of cancer.Scientific Reports, 6(1):24169, 2016
work page 2016
-
[14]
R.L. Mort, R.J.H. Ross, K.J. Hainey, O.J. Harrison, M.A Keighren, G. Landini, R.E. Baker, K.J. Painter, I.J. Jackson, and C.A. Yates. Reconciling diverse mammalian pig- mentation patterns with a fundamental mathematical model.Nature Communications, 7(1):10288, 2016
work page 2016
-
[15]
C. Turner, A.R. Stinchcombe, M. Kohandel, S. Singh, and S. Sivaloganathan. Charac- terization of brain cancer stem cells: a mathematical approach.Cell Proliferation, 42 (4):529–540, 2009
work page 2009
- [16]
-
[17]
A. Golubev. Applications and implications of the exponentially modified gamma distri- bution as a model for time variabilities related to cell proliferation and gene expression. Journal of Theoretical Biology, 393:203–217, 2016
work page 2016
-
[18]
C.A. Yates, M.J. Ford, and R.L. Mort. A Multi-stage Representation of Cell Prolifera- tion as a Markov Process.Bulletin of Mathematical Biology, 79(12):2905–2928, 2017
work page 2017
-
[19]
R.E. Baker and M.J. Simpson. Correcting mean-field approximations for birth-death- movement processes.Physical Review E—Statistical, Nonlinear, and Soft Matter Physics, 82(4):041905, 2010. 52
work page 2010
-
[20]
M.J. Simpson, K.A. Landman, and B.D. Hughes. Multi-species simple exclusion pro- cesses.Physica A: Statistical Mechanics and its Applications, 388(4):399–406, 2009
work page 2009
-
[21]
M.J. Simpson, K.A. Landman, and B.D. Hughes. Cell invasion with proliferation mech- anisms motivated by time-lapse data.Physica A: Statistical Mechanics and its Appli- cations, 389(18):3779–3790, 2010
work page 2010
-
[22]
M.J. Simpson, W. Jin, Sean T. Vittadello, T.A. Tambyah, J.M. Ryan, G. Gunasingh, N.K. Haass, and S.W. McCue. Stochastic models of cell invasion with fluorescent cell cycle indicators.Physica A: Statistical Mechanics and its Applications, 510:375–386, 2018
work page 2018
-
[23]
E. Gavagnin, M.J. Ford, R.L. Mort, T. Rogers, and C.A. Yates. The invasion speed of cell migration models with realistic cell cycle time distributions.Journal of Theoretical Biology, 481:91–99, 2019
work page 2019
-
[24]
K.A. Landman and A.E. Fernando. Myopic random walkers and exclusion processes: Single and multispecies.Physica A: Statistical Mechanics and its Applications, 390 (21-22):3742–3753, 2011
work page 2011
-
[25]
S.T. Vittadello, S.W. McCue, G. Gunasingh, N.K. Haass, and M.J. Simpson. Mathe- matical models incorporating a multi-stage cell cycle replicate normally-hidden inherent synchronization in cell proliferation.Journal of the Royal Society Interface, 16(157), 2019
work page 2019
-
[26]
E. Gavagnin, S.T. Vittadello, G. Gunasingh, M.J. Simpson, N.K. Haass, T. Rogers, and C.A. Yates. Synchronized oscillations in growing cell populations are explained by demographic noise.Biophysical Journal, 120(8):1314–1322, 2021
work page 2021
-
[27]
S.T. Vittadello, S.W. McCue, G. Gunasingh, N.K. Haass, and M.J. Simpson. A novel mathematical model of heterogeneous cell proliferation.Journal of Mathematical Biol- ogy, 82(5):34, 2021
work page 2021
-
[28]
Murray.Mathematical Biology: I
J.D. Murray.Mathematical Biology: I. An Introduction, volume 17. Springer Science & Business Media, 2007
work page 2007
-
[29]
S.T. Vittadello, S.W. McCue, G. Gunasingh, N.K. Haass, and M.J. Simpson. Mathe- matical Models for Cell Migration with Real-Time Cell Cycle Dynamics.Biophysical Journal, 114(5):1241–1253, 2018
work page 2018
-
[30]
C. Falcó, D.J. Cohen, J.A. Carrillo, and R.E. Baker. Quantifying cell cycle regulation by tissue crowding.Biophysical Journal, 124(6):923–932, 2025. 53
work page 2025
-
[31]
M.J. Simpson, B.D. Hughes, and K.A. Landman. Diffusing Populations: Ghosts or Folks?Australasian Journal of Engineering Education, 15(2):59–68, 2009
work page 2009
-
[32]
D.G. Kendall. On the Role of Variable Generation Time in the Development of a Stochastic Birth Process.Biometrika, 35(3/4):316–330, 1948
work page 1948
-
[33]
S.J. Streichan, C.R. Hoerner, T. Schneidt, D. Holzer, and L. Hufnagel. Spatial con- straints control cell proliferation in tissues.Proceedings of the National Academy of Sciences, 111(15):5586–5591, 2014
work page 2014
-
[34]
L. Donker, R. Houtekamer, M. Vliem, F. Sipieter, H. Canever, M. Gómez-González, M. Bosch-Padrós, W.J. Pannekoek, X. Trepat, N. Borghi, and M. Gloerich. A me- chanical G2 checkpoint controls epithelial cell division through E-cadherin-mediated regulation of Wee1-Cdk1.Cell Reports, 41(2):111475, 2022
work page 2022
-
[35]
P.M. Kulesa, C.M. Bailey, J.C. Kasemeier-Kulesa, and R. McLennan. Cranial neural crest migration: New rules for an old road.Developmental Biology, 344(2):543–554, 2010
work page 2010
-
[36]
P.K. Maini, D.L.S. McElwain, and D. Leavesley. Travelling waves in a wound healing assay.Applied Mathematics Letters, 17(5):575–580, 2004
work page 2004
-
[37]
G.C. Paul, Tauhida, and D. Kumar. Revisiting Fisher-KPP model to interpret the spatial spreading of invasive cell population in biology.Heliyon, 8(10):e10773, 2022
work page 2022
-
[38]
D.C. Markham, M.J. Simpson, P.K. Maini, E.A. Gaffney, and R.E. Baker. Comparing methods for modelling spreading cell fronts.Journal of Theoretical Biology, 353:95–103, 2014
work page 2014
- [39]
-
[40]
A. Parker, M.J. Simpson, and R.E. Baker. The impact of experimental design choices on parameter inference for models of growing cell colonies.Royal Society Open Science, 5(8):180384, 2018
work page 2018
-
[41]
E. Gavagnin, J.P. Owen, and C.A. Yates. Pair correlation functions for identifying spatial correlation in discrete domains.Physical Review E, 97(6):062104, 2018
work page 2018
-
[42]
A. Sakaue-Sawano, H. Kurokawa, T. Morimura, A. Hanyu, H. Hama, H. Osawa, S. Kashiwagi, K. Fukami, T. Miyata, H. Miyoshi, and others. Visualizing spatiotemporal dynamics of multicellular cell-cycle progression.Cell, 132(3):487–498, 2008
work page 2008
-
[43]
W. Van Saarloos. Front propagation into unstable states.Physics Reports, 386(2-6): 54 29–222, 2003
work page 2003
-
[44]
J.U. Harrison and C.A Yates. A hybrid algorithm for coupling partial differential equa- tion and compartment-based dynamics.Journal of The Royal Society Interface, 13 (122):20160335, 2016
work page 2016
-
[45]
E. Brunet and B. Derrida. Shift in the velocity of a front due to a cutoff.Physical Review E, 56(3):2597–2604, 1997
work page 1997
-
[46]
D. Selmeczi, S. Mosler, P.H. Hagedorn, N.B. Larsen, and H. Flyvbjerg. Cell motility as persistent random motion: theories from experiments.Biophysical Journal, 89(2): 912–931, 2005
work page 2005
-
[47]
P.H. Wu, A. Giri, S.X. Sun, and D. Wirtz. Three-dimensional cell migration does not follow a random walk.Proceedings of the National Academy of Sciences, 111(11): 3949–3954, 2014
work page 2014
- [48]
-
[49]
Y. Artemenko, T.J. Lampert, and P.N. Devreotes. Moving towards a paradigm: com- mon mechanisms of chemotactic signaling in Dictyostelium and mammalian leukocytes. Cellular and Molecular Life Sciences, 71(19):3711–3747, 2014
work page 2014
-
[50]
J.A. Espina, C.L. Marchant, and E.H. Barriga. Durotaxis: the mechanical control of directed cell migration.The FEBS Journal, 289(10):2736–2754, 2022
work page 2022
-
[51]
S.R. Noureen, R.L. Mort, and C.A. Yates. Modeling adhesion in stochastic and mean- field models of cell migration.Physical Review E, 111(1):014419, 2025. 55
work page 2025
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.