DMFT analysis of Hopfield network with plasticity
Pith reviewed 2026-05-22 02:17 UTC · model grok-4.3
The pith
Moderate plasticity in Hopfield networks enlarges retrieval basins and raises memory capacity via stabilizing delayed feedback.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
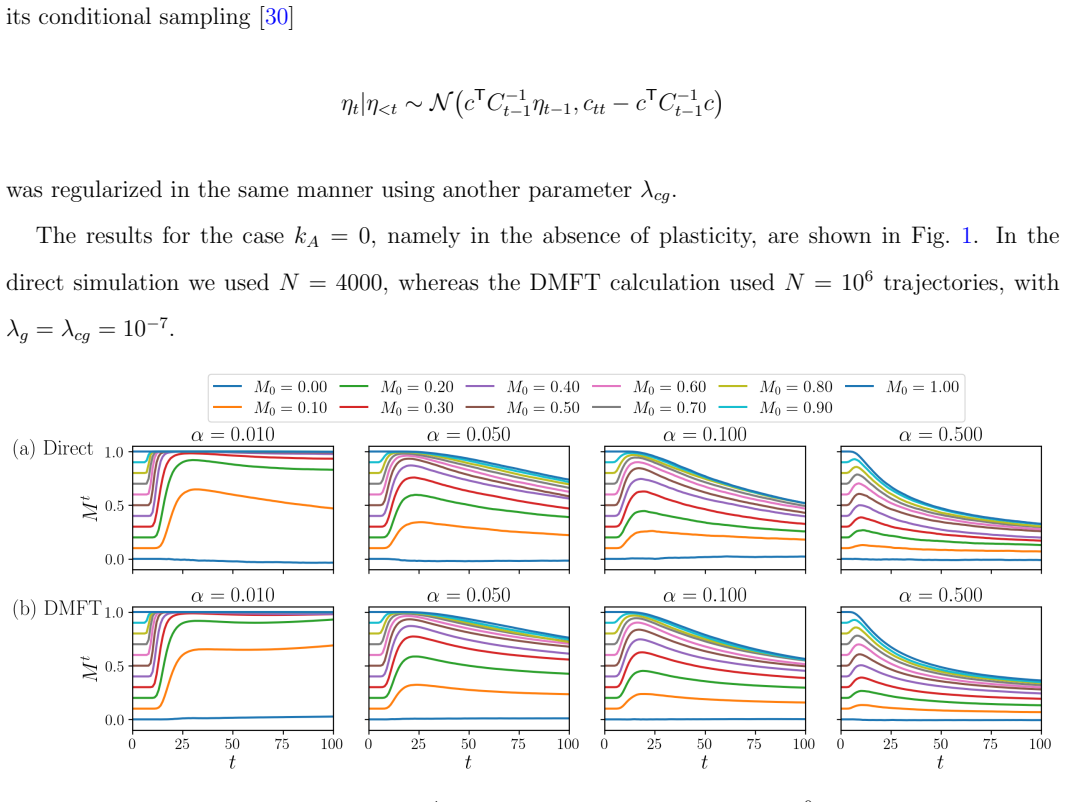
In the large-system limit with many random patterns, the generating-functional approach reduces the joint neural-synaptic dynamics to a single-site stochastic process driven by colored Gaussian crosstalk noise plus delayed feedback from the plasticity rule. Moderate plasticity strength produces positive delayed feedback that stabilizes correct retrieval states against noise, thereby enlarging basins of attraction and increasing the maximum load of retrievable patterns. Stronger plasticity imprints the imperfect initial cue as a fixed point, generating spurious attractors that degrade retrieval quality. An optimal plasticity value therefore emerges from this trade-off.
What carries the argument
The effective single-site stochastic process with colored Gaussian crosstalk noise and delayed feedback terms obtained from dynamical mean-field theory, which tracks the coevolution of neuron states and synaptic weights.
If this is right
- Moderate plasticity increases both the basin volume around each memory and the maximum number of patterns that can be retrieved above the static-network threshold.
- An optimal finite plasticity strength exists that maximizes retrieval performance before cue imprinting dominates.
- The DMFT equations, once solved, quantitatively match the retrieval trajectories seen in direct Monte Carlo simulations of the full network.
- Excessive plasticity strength converts the initial cue into a stable spurious attractor, lowering overall success rate.
Where Pith is reading between the lines
- Biological circuits that use online synaptic changes during recall may operate near this optimal plasticity window to balance stability against overwriting.
- The same delayed-feedback mechanism could be tested in sparse or diluted Hopfield models to check whether the optimum shifts with connectivity.
- Adding a separate slow consolidation phase after retrieval might suppress the spurious attractors created by strong plasticity.
- The single-site reduction supplies a practical way to optimize plasticity rules in artificial associative-memory hardware without simulating every synapse.
Load-bearing premise
The full many-body dynamics can be replaced by an effective single-neuron process whose noise remains Gaussian and colored under the large-system limit with extensively many random patterns.
What would settle it
Direct simulations of finite but large networks in which increasing plasticity strength never produces a peak in basin size or retrieval capacity, but instead shows monotonic degradation, would contradict the predicted optimum.
Figures









read the original abstract
We study a fully connected Hopfield-type associative memory network with online activity-dependent synaptic plasticity, where neural states and synaptic couplings coevolve during retrieval. Using the generating-functional formalism, we derive a dynamical mean-field theory (DMFT) in the large-system limit with extensively many stored random patterns, and show that the many-body dynamics reduces to an effective single-site stochastic process with colored Gaussian crosstalk noise and delayed feedback terms. Numerical solutions of the DMFT equations agree well with direct simulations. We find that moderate plasticity enlarges the basin of attraction and increases the maximum retrievable memory load by generating a positive delayed feedback that stabilizes retrieval against crosstalk noise. However, excessively strong plasticity causes the network to imprint the imperfect initial cue itself, leading to spurious attractors and degraded retrieval performance. Consequently, an optimal plasticity strength emerges from the trade-off between memory stabilization and premature cue imprinting. These results extend the DMFT description of associative memory to networks with coevolving neural and synaptic dynamics.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript applies the generating-functional formalism to derive a dynamical mean-field theory (DMFT) for a fully connected Hopfield associative memory network in which neural states and synaptic weights coevolve under online activity-dependent plasticity. In the N→∞ limit with p=αN random patterns, the many-body dynamics reduces to an effective single-site stochastic process driven by colored Gaussian crosstalk noise plus delayed feedback kernels generated by the plasticity rule. Numerical integration of the resulting DMFT equations is reported to agree with direct simulations. The central finding is that moderate plasticity enlarges the basin of attraction and raises the maximum retrievable load α_c by producing a positive delayed feedback that counters crosstalk, whereas excessively strong plasticity imprints the imperfect initial cue, creating spurious attractors and degrading performance; an optimal plasticity strength therefore emerges from this trade-off.
Significance. If the DMFT closure and the reported numerical agreement hold, the work supplies a controlled theoretical extension of mean-field methods to coevolving neural-synaptic dynamics, furnishing an explicit mechanism (delayed positive feedback) for the stabilizing effect of moderate plasticity and a concrete explanation for the existence of an optimal regime. The explicit large-N derivation and the quantitative match to simulations constitute reproducible, falsifiable content that strengthens the result relative to purely phenomenological models.
major comments (2)
- [§2] §2 (DMFT derivation): the self-consistent closure for the delayed-feedback kernel K(t,t′) is expressed in terms of the same two-time correlation functions that are themselves determined by the single-site process; while this is standard in generating-functional DMFT, the manuscript does not provide an explicit check that the Gaussianity assumption for the effective noise remains valid when the plasticity strength pushes the system near the imprinting transition (where cue-specific correlations may become non-negligible).
- [Results section] Results section (comparison with simulations): the statement that DMFT solutions “agree well” with direct simulations is not accompanied by quantitative error measures (e.g., integrated squared difference on overlap trajectories), the range of α and plasticity strengths tested, or the finite-N values employed; without these, the support for the claimed optimal-plasticity trade-off remains only moderately quantitative.
minor comments (2)
- [Notation] Notation: the definition of the plasticity strength parameter (denoted variously as λ or η in different paragraphs) should be unified and placed in a single equation early in the text.
- [Figures] Figure captions: the legends for the overlap-vs-time curves do not explicitly state the initial cue overlap m(0) used in each panel, making it difficult to reproduce the basin-enlargement claim.
Simulated Author's Rebuttal
We thank the referee for the careful reading of our manuscript and the constructive comments. We address each major comment below and outline the revisions we will make to strengthen the presentation.
read point-by-point responses
-
Referee: [§2] §2 (DMFT derivation): the self-consistent closure for the delayed-feedback kernel K(t,t′) is expressed in terms of the same two-time correlation functions that are themselves determined by the single-site process; while this is standard in generating-functional DMFT, the manuscript does not provide an explicit check that the Gaussianity assumption for the effective noise remains valid when the plasticity strength pushes the system near the imprinting transition (where cue-specific correlations may become non-negligible).
Authors: We agree that an explicit verification of the Gaussianity assumption would be useful near the imprinting transition. Although the central-limit argument for Gaussian crosstalk noise holds formally in the N→∞ limit, cue-specific correlations could in principle become relevant when plasticity is strong. In the revised manuscript we will add a short subsection with a numerical check: we extract the effective noise distribution from finite-N simulations at plasticity strengths approaching the transition and compare its kurtosis to the Gaussian value of 3. This will either confirm the assumption or identify the parameter region where deviations appear. revision: yes
-
Referee: [Results section] Results section (comparison with simulations): the statement that DMFT solutions “agree well” with direct simulations is not accompanied by quantitative error measures (e.g., integrated squared difference on overlap trajectories), the range of α and plasticity strengths tested, or the finite-N values employed; without these, the support for the claimed optimal-plasticity trade-off remains only moderately quantitative.
Authors: We accept that the current comparison lacks quantitative detail. In the revised manuscript we will report (i) integrated squared differences between DMFT and simulated overlap trajectories, (ii) the explicit ranges of memory load α and plasticity strength examined, and (iii) the finite system sizes N used (typically N ≥ 1000). These additions will make the quantitative support for the optimal-plasticity regime more rigorous. revision: yes
Circularity Check
DMFT derivation is self-contained via generating-functional formalism and validated by simulations
full rationale
The paper derives its DMFT equations from the generating-functional formalism applied to the large-N limit with extensively many random patterns, reducing the many-body dynamics to an effective single-site stochastic process. This reduction is obtained directly from the formalism rather than by fitting or self-definition, and the resulting self-consistent equations for noise and feedback kernels are solved numerically with quantitative agreement to direct simulations. No load-bearing step reduces by construction to its own inputs, no fitted parameter is renamed as a prediction, and no uniqueness theorem or ansatz is imported via self-citation in a way that forces the central claims. The trade-off between moderate and excessive plasticity follows from the structure of the independently derived equations.
Axiom & Free-Parameter Ledger
free parameters (1)
- plasticity strength
axioms (2)
- domain assumption Large-system limit with extensively many stored random patterns
- domain assumption Generating-functional formalism applies to the joint neural-synaptic dynamics
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
the many-body dynamics reduces to an effective single-site stochastic process with colored Gaussian crosstalk noise and delayed feedback terms
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Neural networks and physical systems with emergent collective computational abilities
J J Hopfield. Neural networks and physical systems with emergent collective computational abilities. Proceedings of the National Academy of Sciences, 79(8):2554–2558, 1982
work page 1982
-
[2]
S-i Amari, Learning patterns and pattern sequences by self-organizing nets of threshold elements.IEEE Transactions on computers, 100(11):1197–1206, 1972
work page 1972
-
[3]
Ludovica Serricchio, Dario Bocchi, Claudio Chilin, Raffaele Marino, Matteo Negri, Chiara Cammarota, and Federico Ricci-Tersenghi. Daydreaming hopfield networks and their surprising effectiveness on correlated data.Neural Networks, 186:107216, 2025. 33
work page 2025
-
[4]
Daydreaming algorithm for biased patterns, 2026
Mikiya Doi, Masayuki Ohzeki, and Federico Ricci-Tersenghi. Daydreaming algorithm for biased patterns, 2026
work page 2026
-
[5]
Analysis of the hopfield model incorpo- rating the effects of unlearning, 2026
Shuta Takeuchi, Takashi Takahashi, and Yoshiyuki Kabashima. Analysis of the hopfield model incorpo- rating the effects of unlearning, 2026
work page 2026
-
[6]
Daniel J. Amit, Hanoch Gutfreund, and H. Sompolinsky. Storing infinite numbers of patterns in a spin-glass model of neural networks.Phys. Rev. Lett., 55:1530–1533, Sep 1985
work page 1985
-
[7]
Guy Major and David Tank. Persistent neural activity: prevalence and mechanisms.Current Opinion in Neurobiology, 14(6):675–684, 2004
work page 2004
-
[8]
Misha V. Tsodyks and Henry Markram. The neural code between neocortical pyramidal neurons depends on neurotransmitter release probability.Proceedings of the National Academy of Sciences, 94(2):719–723, 1997
work page 1997
-
[9]
Guo-qiang Bi and Mu-ming Poo. Synaptic modifications in cultured hippocampal neurons: Dependence on spike timing, synaptic strength, and postsynaptic cell type.Journal of Neuroscience, 18(24):10464–10472, 1998
work page 1998
-
[10]
Short-term synaptic plasticity.Annual review of physiology, 64:355–405, 02 2002
Robert Zucker and Wade Regehr. Short-term synaptic plasticity.Annual review of physiology, 64:355–405, 02 2002
work page 2002
-
[11]
Synaptic theory of working memory.Science, 319(5869):1543–1546, 2008
Gianluigi Mongillo, Omri Barak, and Misha Tsodyks. Synaptic theory of working memory.Science, 319(5869):1543–1546, 2008
work page 2008
-
[12]
Katie C. Bittner, Aaron D. Milstein, Christine Grienberger, Sandro Romani, and Jeffrey C. Magee. Be- havioral time scale synaptic plasticity underlies ca1 place fields.Science, 357(6355):1033–1036, 2017
work page 2017
-
[13]
Aaron D Milstein, Yiding Li, Katie C Bittner, Christine Grienberger, Ivan Soltesz, Jeffrey C Magee, and Sandro Romani. Bidirectional synaptic plasticity rapidly modifies hippocampal representations.eLife, 10:e73046, dec 2021
work page 2021
-
[14]
Neural networks with dynamic synapses.Neural Computation, 10:821–835, 05 1998
Michail Tsodyks, Klaus Pawelzik, and Henry Markram. Neural networks with dynamic synapses.Neural Computation, 10:821–835, 05 1998
work page 1998
-
[15]
Associative memory with dynamic synapses.Neural Computation, 14:2903–2923, 12 2002
Lovorka Pantic, Joaqu´ ın Torres, Hilbert Kappen, and Stan Gielen. Associative memory with dynamic synapses.Neural Computation, 14:2903–2923, 12 2002
work page 2002
-
[16]
Jorge F. Mejias and Joaqu´ ın J. Torres. Maximum memory capacity on neural networks with short-term synaptic depression and facilitation.Neural Computation, 21(3):851–871, 03 2009. 34
work page 2009
-
[17]
Yosuke Otsubo, Kenji Nagata, Masafumi Oizumi, and Masato Okada. Instabilities in associative memory model with synaptic depression and switching phenomena among attractors.Journal of the Physical Society of Japan, 79, 05 2010
work page 2010
-
[18]
Yuichi Katori, Yosuke Otsubo, Masato Okada, and Kazuyuki Aihara. Stability analysis of associative memory network composed of stochastic neurons and dynamic synapses.Frontiers in Computational Neuroscience, Volume 7 - 2013, 2013
work page 2013
-
[19]
Yasuhiko Igarashi, Masafumi Oizumi, and Masato Okada. Mean field analysis of stochastic neural network models with synaptic depression.Journal of the Physical Society of Japan, 79, 03 2010
work page 2010
-
[20]
David G. Clark and L. F. Abbott. Theory of coupled neuronal-synaptic dynamics.Phys. Rev. X, 14:021001, Apr 2024
work page 2024
-
[21]
Yoshiyuki Kabashima and Kazushi Mimura. Dynamical mean field approach to associative memory model with non-monotonic transfer functions.Journal of Statistical Mechanics: Theory and Experiment, 2026(1):014002, jan 2026
work page 2026
-
[22]
H. Sompolinsky, A. Crisanti, and H. J. Sommers. Chaos in random neural networks.Phys. Rev. Lett., 61:259–262, Jul 1988
work page 1988
-
[23]
P. C. Martin, E. D. Siggia, and H. A. Rose. Statistical dynamics of classical systems.Phys. Rev. A, 8:423–437, Jul 1973
work page 1973
-
[24]
C. De Dominicis. Dynamics as a substitute for replicas in systems with quenched random impurities.Phys. Rev. B, 18:4913–4919, Nov 1978
work page 1978
-
[25]
John A Hertz, Yasser Roudi, and Peter Sollich. Path integral methods for the dynamics of stochastic and disordered systems.Journal of Physics A: Mathematical and Theoretical, 50(3):033001, dec 2016
work page 2016
-
[26]
Wenxuan Zou and Haiping Huang. Introduction to dynamical mean-field theory of randomly connected neural networks with bidirectionally correlated couplings.SciPost Phys. Lect. Notes, page 79, 2024
work page 2024
-
[27]
H. Eissfeller and M. Opper. New method for studying the dynamics of disordered spin systems without finite-size effects.Phys. Rev. Lett., 68:2094–2097, Mar 1992
work page 2094
-
[28]
Arthur E. Hoerl and Robert W. Kennard. Ridge regression: Biased estimation for nonorthogonal problems. Technometrics, 12(1):55–67, 1970
work page 1970
-
[29]
A well-conditioned estimator for large-dimensional covariance matrices
Olivier Ledoit and Michael Wolf. A well-conditioned estimator for large-dimensional covariance matrices. Journal of Multivariate Analysis, 88(2):365–411, February 2004. 35
work page 2004
-
[30]
Carl Edward Rasmussen and Christopher K. I. Williams.Gaussian Processes for Machine Learning. The MIT Press, 11 2005
work page 2005
-
[31]
Daiki Goto, Hector Rios, Monika Scholz, and Suriyanarayanan Vaikuntanathan. Neuromodulation-inspired gated associative memory networks:extended memory retrieval and emergent multistability.ArXiv, 12 2025
work page 2025
-
[32]
Albert J. Wakhloo, David G. Clark, and L.F. Abbott. Associative synaptic plasticity creates dynamic persistent activity.bioRxiv, 2025. 36
work page 2025
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.