Predictions for and lack of maximal information transmission in the neuromuscular junction
Pith reviewed 2026-06-27 07:18 UTC · model grok-4.3
The pith
The Drosophila neuromuscular junction does not maximize information transmission from nerve to muscle.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
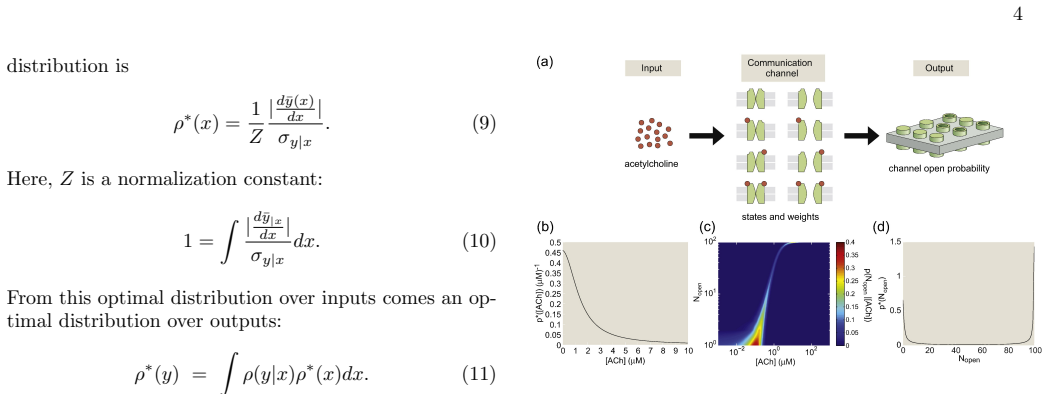
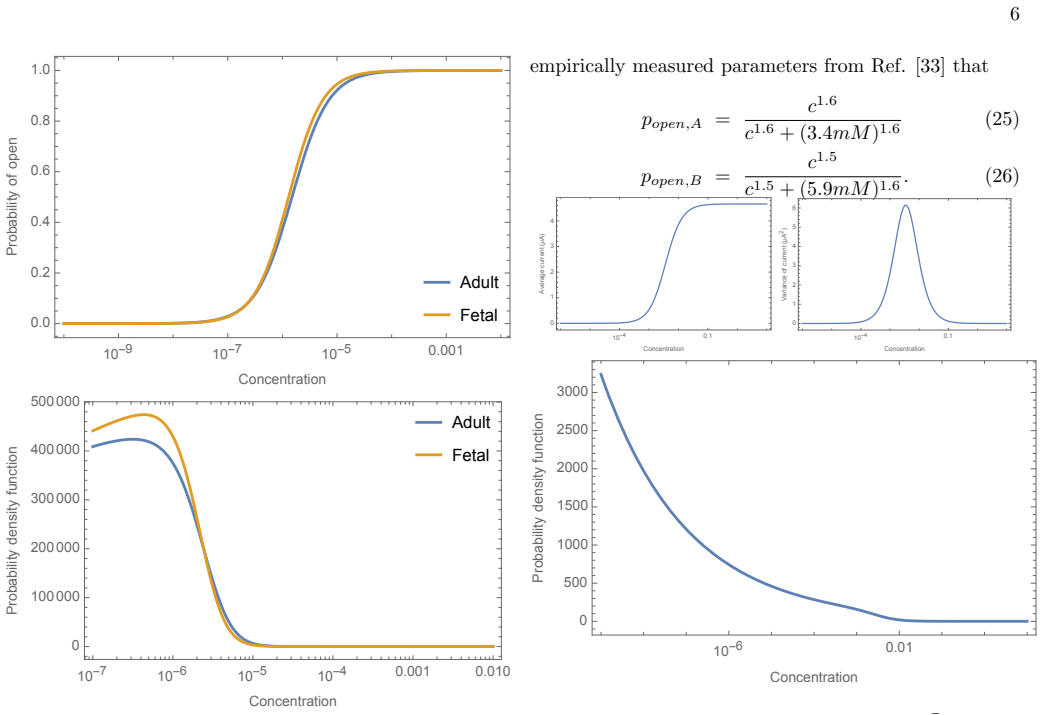
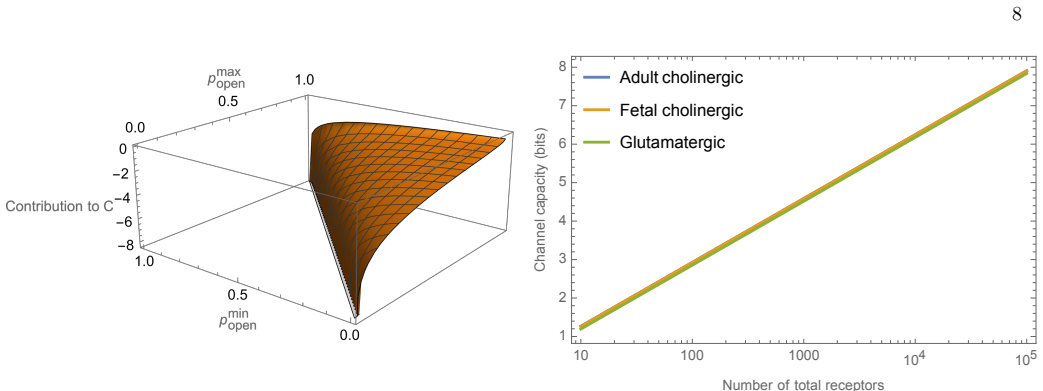
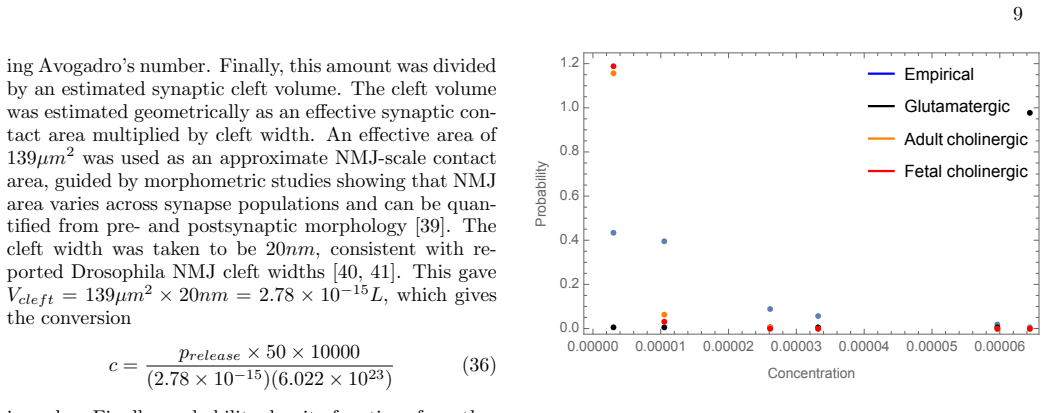
An information maximization analysis applied to the transformation from neurotransmitter concentration to current at cholinergic and glutamatergic neuromuscular junctions produces a theoretical distribution over concentrations. Comparison of this distribution to the experimentally measured distribution of synaptic vesicle release probabilities at the Drosophila neuromuscular junction reveals very little agreement. This indicates that the Drosophila NMJ does not shape its distribution of synaptic vesicle release probabilities in order to maximize information transmission from nervous system to muscle. The analysis supplies explicit predictions for cholinergic systems.
What carries the argument
Information maximization analysis applied to biological dose-response relationships, which computes the distribution over neurotransmitter concentrations that maximizes transmitted information.
If this is right
- The Drosophila NMJ release probability distribution deviates substantially from the information-maximizing distribution derived from dose-response curves.
- Cholinergic neuromuscular junctions have explicit predicted optimal distributions that differ from the glutamatergic case.
- Information transmission is not the dominant constraint that determines the observed distribution of release probabilities at the Drosophila NMJ.
Where Pith is reading between the lines
- The mismatch suggests that other functional demands, such as energy use or robustness, may dominate the shaping of release probabilities instead.
- The same information-maximization procedure could be applied to other synapses to test whether maximization occurs outside the NMJ.
- If the conclusion is general, models of neural coding at peripheral junctions will need to incorporate non-information criteria from the start.
Load-bearing premise
That the information maximization analysis produces the correct optimal distribution over neurotransmitter concentrations when based on the biological dose-response relationships.
What would settle it
A new experimental measurement of the distribution of synaptic vesicle release probabilities at the Drosophila NMJ that closely matches the theoretically derived optimal distribution would falsify the claim of no maximization.
Figures

read the original abstract
A key question in theoretical biology is how effectively biological systems preserve information about their inputs while operating under physical and functional constraints. We examine that question at the neuromuscular junction (NMJ) by studying how neurotransmitter concentration is transformed into current at both cholinergic and glutamatergic NMJs. An information maximization analysis was used to derive a theoretical distribution over neurotransmitter concentrations based on biological understandings of dose-response relationships. These theoretical distributions were compared to an experimentally derived distribution obtained from a Drosophila NMJ. The theoretical and experimental distributions showed very little agreement, indicating that the Drosophila NMJ does not shape its distribution of synaptic vesicle release probabilities in order to maximize information transmission from nervous system to muscle. Predictions for cholinergic systems are provided.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript derives an information-maximizing distribution over neurotransmitter concentrations at NMJs from biological dose-response curves for both cholinergic and glutamatergic synapses. It then compares this theoretical distribution to an experimentally measured distribution from the Drosophila NMJ and reports very little agreement, concluding that the NMJ does not shape its synaptic vesicle release probability distribution to maximize information transmission. Predictions for cholinergic systems are also provided.
Significance. If the comparison between the derived optimal distribution and the experimental data is valid, the result would indicate that information maximization is not a governing principle for release statistics at the NMJ. This would be a substantive contribution to theoretical biology on optimality constraints in synaptic transmission. The explicit predictions for cholinergic systems constitute a falsifiable output that strengthens the work.
major comments (1)
- [Abstract] Abstract: The information-maximization analysis produces a theoretical distribution over neurotransmitter concentrations, yet the central claim and the experimental comparison concern the distribution of synaptic vesicle release probabilities. Release probability determines concentration only through a nonlinear, stochastic mapping that depends on vesicle content, cleft geometry, and diffusion; without an explicit transformation step that converts one distribution into the other before comparison, the reported mismatch does not directly falsify the maximization hypothesis for release probabilities.
Simulated Author's Rebuttal
We thank the referee for their careful reading of the manuscript and for identifying an important distinction between the theoretical and experimental quantities. We address the major comment below and outline the revisions we will make.
read point-by-point responses
-
Referee: [Abstract] Abstract: The information-maximization analysis produces a theoretical distribution over neurotransmitter concentrations, yet the central claim and the experimental comparison concern the distribution of synaptic vesicle release probabilities. Release probability determines concentration only through a nonlinear, stochastic mapping that depends on vesicle content, cleft geometry, and diffusion; without an explicit transformation step that converts one distribution into the other before comparison, the reported mismatch does not directly falsify the maximization hypothesis for release probabilities.
Authors: We agree that the referee correctly identifies a gap in the current presentation. The information-maximization procedure yields an optimal distribution over postsynaptic neurotransmitter concentrations, while the Drosophila data consist of measured vesicle release probabilities. Although the abstract and discussion equate the two for the purpose of the central claim, no explicit mapping (accounting for binomial release, vesicle content variability, and cleft diffusion) is provided. We will revise the manuscript by adding a dedicated section that derives the concentration distribution from the release-probability distribution under a standard synaptic cleft model. The abstract and conclusions will be updated to state the comparison more precisely. This change will make the falsification argument rigorous. revision: yes
Circularity Check
No significant circularity in derivation chain
full rationale
The paper derives a theoretical distribution over neurotransmitter concentrations via information maximization applied to dose-response relationships drawn from biological understandings, then compares the result to a separate experimentally derived distribution. No load-bearing step reduces the claimed prediction to fitted inputs by construction, nor does any self-citation chain or ansatz smuggling appear in the provided text. The central claim rests on an observed mismatch between independently obtained objects; while the skeptic correctly notes that release-probability data and concentration distributions are not identical without an explicit mapping, this is a question of evidential support rather than circularity. The derivation remains self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
From Ref
The approximate cholinergic information channel For cholinergic systems, acetylcholine floods the synapse through release of synaptic vesicles, and this acetylcholine attaches to nAChRs. From Ref. [32], their dose-response relationship when modeled phenomenolog- ically by a Hill model is popen,nAChR = cnH K nH d +c nH (13) If we assume that the amount of ...
-
[2]
Letcbe the concentration of glutamate
The approximate glutamatergic information channel The glutamatergic information channel is slightly more complicated than the cholinergic information channel, simply because there are two slightly different iGluRs that both play an important role in determining the amount of current that flows through the muscle fiber. Letcbe the concentration of glutamat...
2000
-
[3]
C. E. Shannon, A mathematical theory of communica- tion, The Bell system technical journal27, 379 (1948)
1948
-
[4]
Waltermann and E
C. Waltermann and E. Klipp, Information theory based approaches to cellular signaling, Biochimica et Biophys- ica Acta (BBA)-General Subjects1810, 924 (2011)
2011
-
[5]
H. B. Barlowet al., Possible principles underlying the transformation of sensory messages, Sensory communi- cation1, 217 (1961)
1961
-
[6]
S. P. Strong, R. Koberle, R. R. D. R. Van Steveninck, and W. Bialek, Entropy and information in neural spike trains, Physical review letters80, 197 (1998)
1998
-
[7]
Bell and T
A. Bell and T. J. Sejnowski, Edges are the’independent components’ of natural scenes., Advances in neural infor- mation processing systems9(1996)
1996
-
[8]
independent compo- nents
A. J. Bell and T. J. Sejnowski, The “independent compo- nents” of natural scenes are edge filters, Vision research 37, 3327 (1997)
1997
-
[9]
B. A. Olshausen and D. J. Field, Sparse coding of sensory inputs, Current opinion in neurobiology14, 481 (2004)
2004
-
[10]
Tkaˇ cik, A
G. Tkaˇ cik, A. M. Walczak, and W. Bialek, Optimizing information flow in small genetic networks, Physical Re- view E—Statistical, Nonlinear, and Soft Matter Physics 80, 031920 (2009)
2009
-
[11]
Tkaˇ cik, C
G. Tkaˇ cik, C. G. Callan Jr, and W. Bialek, Information flow and optimization in transcriptional regulation, Pro- ceedings of the National Academy of Sciences105, 12265 (2008)
2008
-
[12]
G. Lan, P. Sartori, S. Neumann, V. Sourjik, and Y. Tu, The energy–speed–accuracy trade-off in sensory adapta- tion, Nature physics8, 422 (2012)
2012
-
[13]
Klinger and G
J. Klinger and G. M. Rotskoff, Universal energy-speed- accuracy trade-offs in driven nonequilibrium systems, Physical Review E111, 014114 (2025)
2025
-
[14]
D. H. Wolpert, J. Korbel, C. W. Lynn, F. Tasnim, J. A. Grochow, G. Karde¸ s, J. B. Aimone, V. Balasubramanian, E. De Giuli, D. Doty,et al., Is stochastic thermodynamics the key to understanding the energy costs of computa- tion?, Proceedings of the National Academy of Sciences 121, e2321112121 (2024)
2024
-
[15]
B. M. Martins and P. S. Swain, Trade-offs and con- straints in allosteric sensing, Plos computational biology 7, e1002261 (2011)
2011
-
[16]
R. Bhui, L. Lai, and S. J. Gershman, Resource-rational decision making, Current Opinion in Behavioral Sciences 41, 15 (2021)
2021
-
[17]
Lieder and T
F. Lieder and T. L. Griffiths, Resource-rational analysis: Understanding human cognition as the optimal use of limited computational resources, Behavioral and brain sciences43, e1 (2020)
2020
-
[18]
A. J. Tjalma, V. Galstyan, J. Goedhart, L. Slim, N. B. Becker, and P. R. Ten Wolde, Trade-offs between cost and information in cellular prediction, Proceedings of the National Academy of Sciences120, e2303078120 (2023)
2023
-
[19]
Laughlinet al., A simple coding procedure enhances a neuron’s information capacity, Z
S. Laughlinet al., A simple coding procedure enhances a neuron’s information capacity, Z. Naturforsch36, 51 (1981)
1981
-
[20]
Duran-Urriago and S
A. Duran-Urriago and S. Marzen, Not so optimal: The evolution of mutual information in potassium voltage- gated channels, Plos one18, e0264424 (2023)
2023
-
[21]
Bouzat and N
C. Bouzat and N. Mukhtasimova, The nicotinic acetyl- choline receptor as a molecular machine for neuromus- cular transmission, Current Opinion in Physiology4, 40 (2018)
2018
-
[22]
E. S. Peled and E. Y. Isacoff, Optical quantal analysis of synaptic transmission in wild-type and rab3-mutant drosophila motor axons, Nature neuroscience14, 519 (2011)
2011
-
[23]
T. M. Cover,Elements of information theory(John Wi- ley & Sons, 1999)
1999
-
[24]
J. B. Kinney and G. S. Atwal, Equitability, mutual in- formation, and the maximal information coefficient, Pro- ceedings of the National Academy of Sciences111, 3354 (2014)
2014
-
[25]
S. E. Marzen and S. DeDeo, The evolution of lossy compression, Journal of The Royal Society Interface14 (2017)
2017
-
[26]
Marzen, Resource-rational reinforcement learning and sensorimotor causal states, and resource-rational max- iminers, Interface Focus15(2025)
S. Marzen, Resource-rational reinforcement learning and sensorimotor causal states, and resource-rational max- iminers, Interface Focus15(2025)
2025
-
[27]
Bialek,Biophysics: searching for principles(Prince- ton University Press, 2012)
W. Bialek,Biophysics: searching for principles(Prince- ton University Press, 2012)
2012
-
[28]
Tostevin and P
F. Tostevin and P. R. Ten Wolde, Mutual information be- tween input and output trajectories of biochemical net- works, Physical review letters102, 218101 (2009)
2009
-
[29]
Cheong, A
R. Cheong, A. Rhee, C. J. Wang, I. Nemenman, and A. Levchenko, Information transduction capacity of noisy biochemical signaling networks, science334, 354 (2011)
2011
-
[30]
N. Unwin, Nicotinic acetylcholine receptor and the struc- tural basis of neuromuscular transmission: insights from torpedo postsynaptic membranes, Quarterly reviews of biophysics46, 283 (2013)
2013
-
[31]
Karlin, Emerging structure of the nicotinic acetyl- choline receptors, Nature Reviews Neuroscience3, 102 (2002)
A. Karlin, Emerging structure of the nicotinic acetyl- choline receptors, Nature Reviews Neuroscience3, 102 (2002)
2002
-
[32]
H. L. Atwood and S. Karunanithi, Diversification of synaptic strength: presynaptic elements, Nature Reviews Neuroscience3, 497 (2002)
2002
-
[33]
Marzen, H
S. Marzen, H. G. Garcia, and R. Phillips, Statistical me- chanics of monod–wyman–changeux (mwc) models, Jour- nal of molecular biology425, 1433 (2013)
2013
-
[34]
R. J. Prince and S. M. Sine, Acetylcholine and epibati- dine binding to muscle acetylcholine receptors distinguish 11 between concerted and uncoupled models, Journal of Bi- ological Chemistry274, 19623 (1999)
1999
-
[35]
T. H. Han, P. Dharkar, M. L. Mayer, and M. Serpe, Func- tional reconstitution of drosophila melanogaster nmj glu- tamate receptors, Proceedings of the National Academy of Sciences112, 6182 (2015)
2015
-
[36]
Phillips, J
R. Phillips, J. Kondev, J. Theriot, and H. Garcia,Phys- ical biology of the cell(Garland Science, 2012)
2012
-
[37]
Soriano and S
J. Soriano and S. Marzen, How well can we infer selec- tion benefits and mutation rates from allele frequencies?, Entropy25, 615 (2023)
2023
-
[38]
Cohen-Saidon, A
C. Cohen-Saidon, A. A. Cohen, A. Sigal, Y. Liron, and U. Alon, Dynamics and variability of erk2 response to egf in individual living cells, Molecular cell36, 885 (2009)
2009
-
[39]
X. R. Bao, I. D. Fraser, E. A. Wall, S. R. Quake, and M. I. Simon, Variability in g-protein-coupled signaling studied with microfluidic devices, Biophysical journal99, 2414 (2010)
2010
- [40]
-
[41]
R. A. Jones, C. D. Reich, K. N. Dissanayake, F. Krist- mundsdottir, G. S. Findlater, R. R. Ribchester, M. W. Simmen, and T. H. Gillingwater, Nmj-morph reveals principal components of synaptic morphology influenc- ing structure–function relationships at the neuromuscu- lar junction, Open Biology6(2016)
2016
-
[42]
Prokop, M
A. Prokop, M. Landgraf, E. Rushton, K. Broadie, and M. Bate, Presynaptic development at the drosophila neu- romuscular junction: assembly and localization of presy- naptic active zones, Neuron17, 617 (1996)
1996
-
[43]
Koper, A
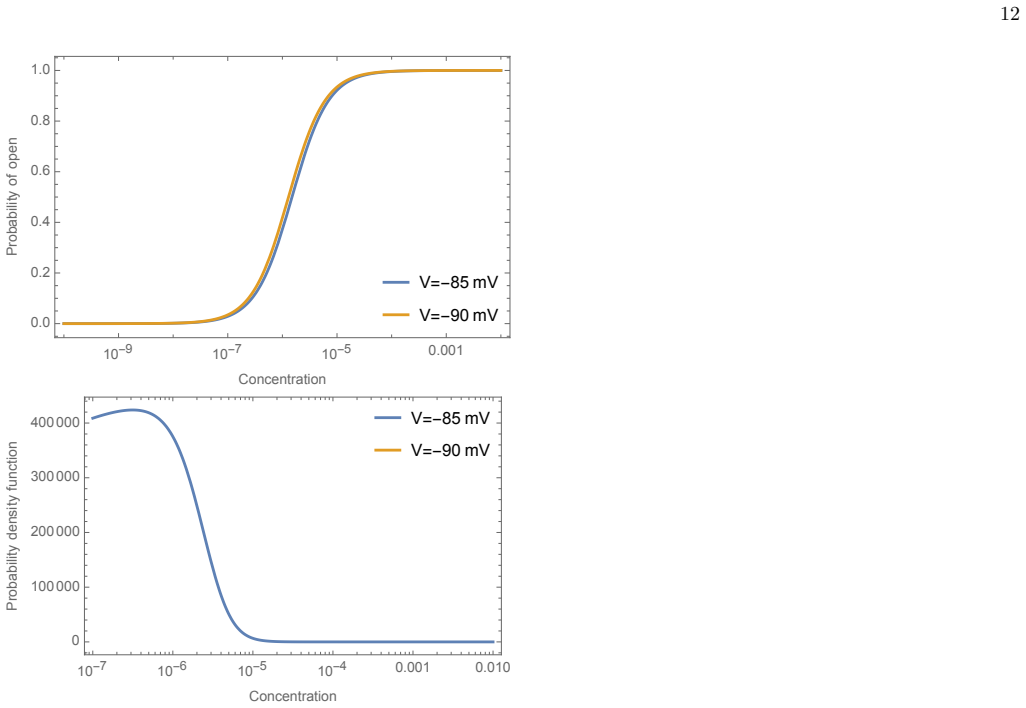
A. Koper, A. Schenck, and A. Prokop, Analysis of adhe- sion molecules and basement membrane contributions to synaptic adhesion at the drosophila embryonic nmj, PloS one7, e36339 (2012). Appendix A: Alterations if equilibrium membrane potential changes As described in Ref. [25], in theory if statistical me- chanics is used, open probabilities of ion channe...
2012
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.