Communication Modeling of Long-Distance Abscisic Acid Signaling in Plant Vascular Systems
Pith reviewed 2026-06-27 02:08 UTC · model grok-4.3
The pith
ABA transport through plant xylem can be modeled as molecular communication where higher release quantities and larger receivers improve signal reception.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
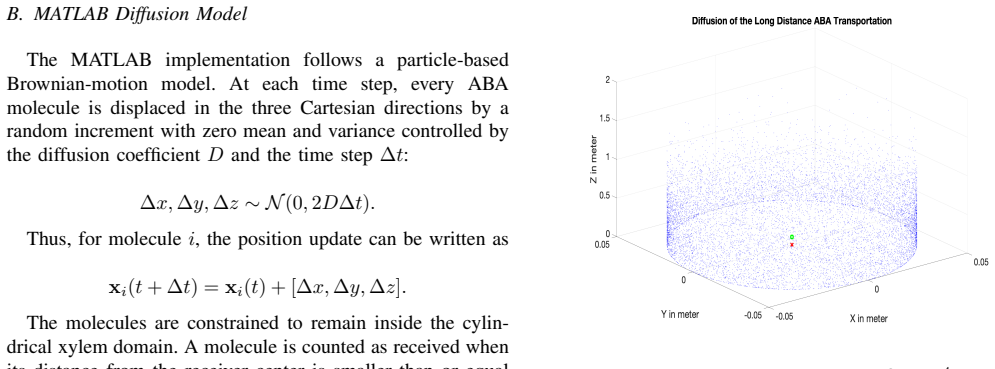
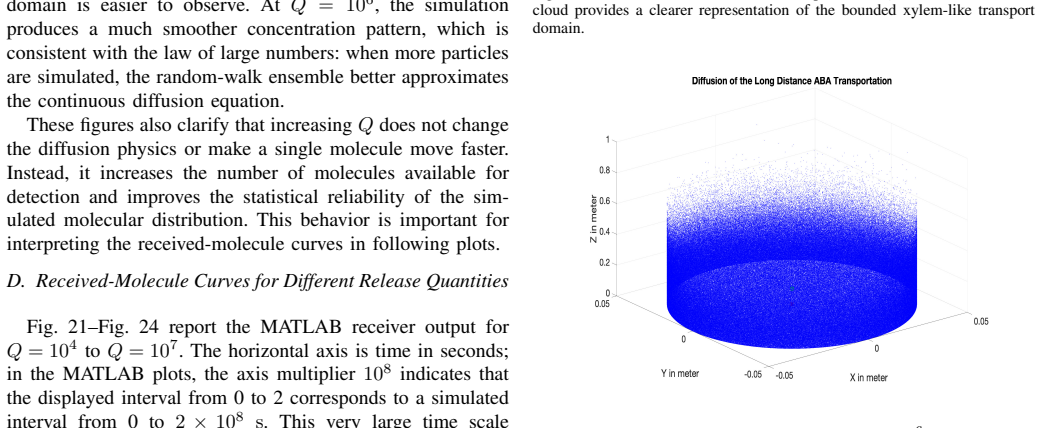
The paper presents a molecular-communication-inspired model of ABA transport in which root-side ABA release is represented as a transmitter, the xylem pathway as a bounded channel, and soybean tissue as a receiver; MATLAB Brownian-motion simulations demonstrate that higher release quantities produce smoother and stronger reception trends while larger receivers increase molecule-capture probability.
What carries the argument
Molecular-communication model treating the xylem as a bounded channel and soybean tissue as receiver in Brownian-motion simulations.
If this is right
- Higher quantities of released ABA molecules produce smoother and stronger reception trends at the receiver.
- Larger receiver radii increase the probability of capturing ABA molecules.
- The simulation framework allows evaluation of how different parameters affect ABA signal detection without requiring new biological experiments for each case.
- The model can be used to explore coordination of plant responses to drought and other stresses via long-distance vascular signaling.
Where Pith is reading between the lines
- The same modeling structure could be applied to other mobile plant hormones to compare their transport efficiencies.
- If the simulated trends hold in living plants, the model might help identify optimal release sites or tissue properties for enhancing stress signaling.
- Incorporating active flow or binding kinetics into the channel description would test whether the passive Brownian assumption remains sufficient.
Load-bearing premise
The xylem pathway can be represented as a bounded channel and soybean tissue as a receiver in a Brownian-motion molecular-communication model that meaningfully reflects real long-distance ABA transport dynamics.
What would settle it
Direct measurements of ABA arrival rates and concentrations in soybean xylem tissue under controlled release conditions that fail to match the simulated trends for varying molecule quantities and receiver sizes.
Figures







read the original abstract
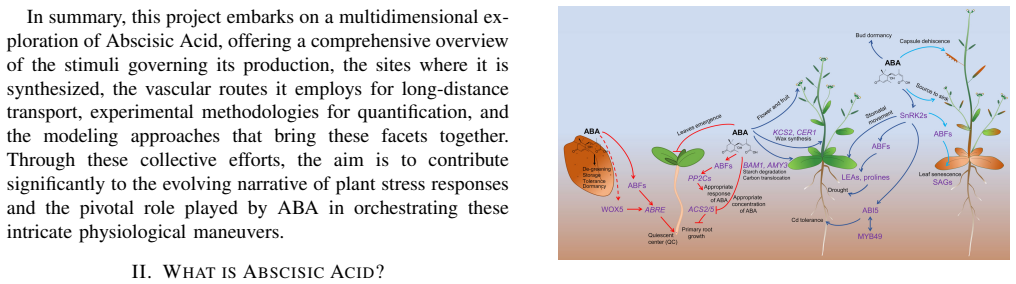
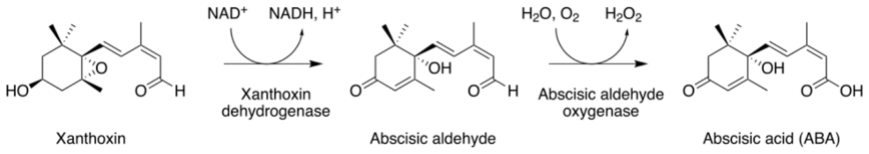
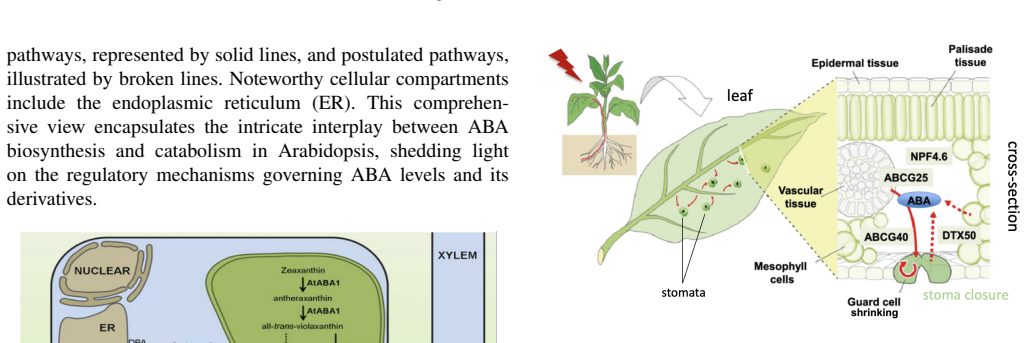
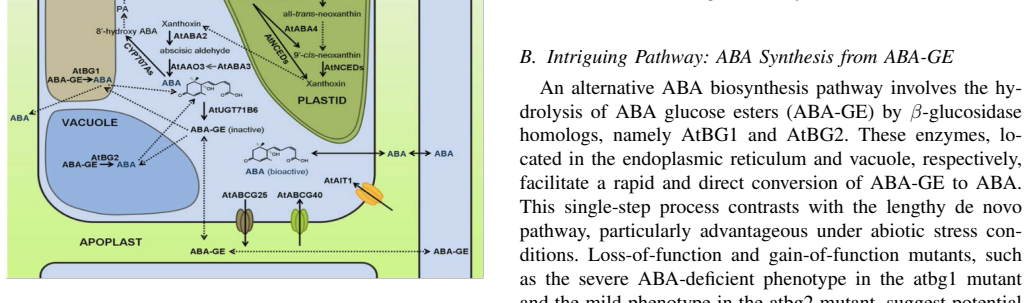
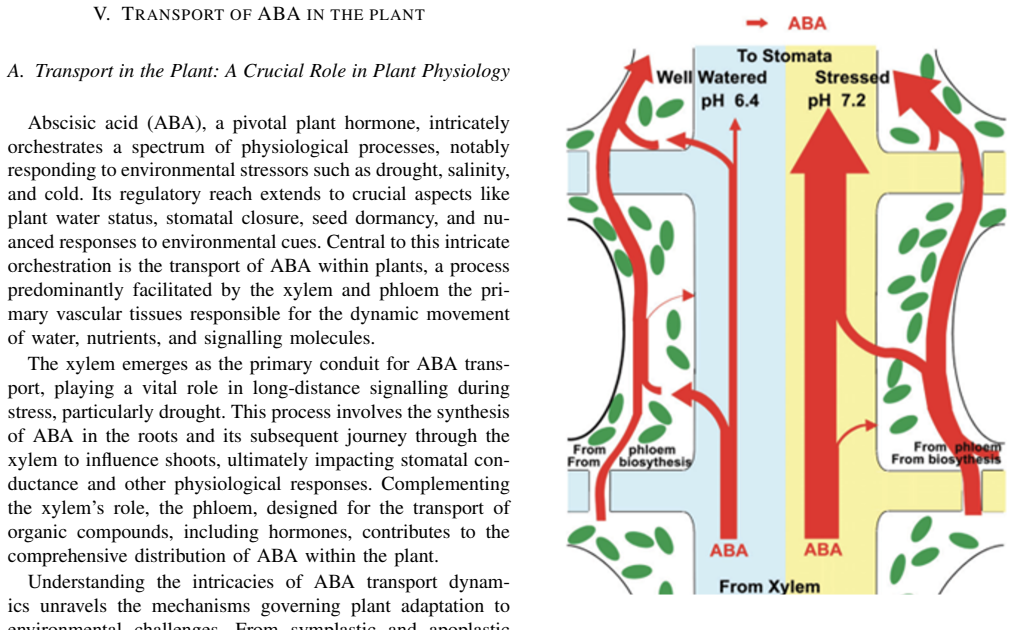
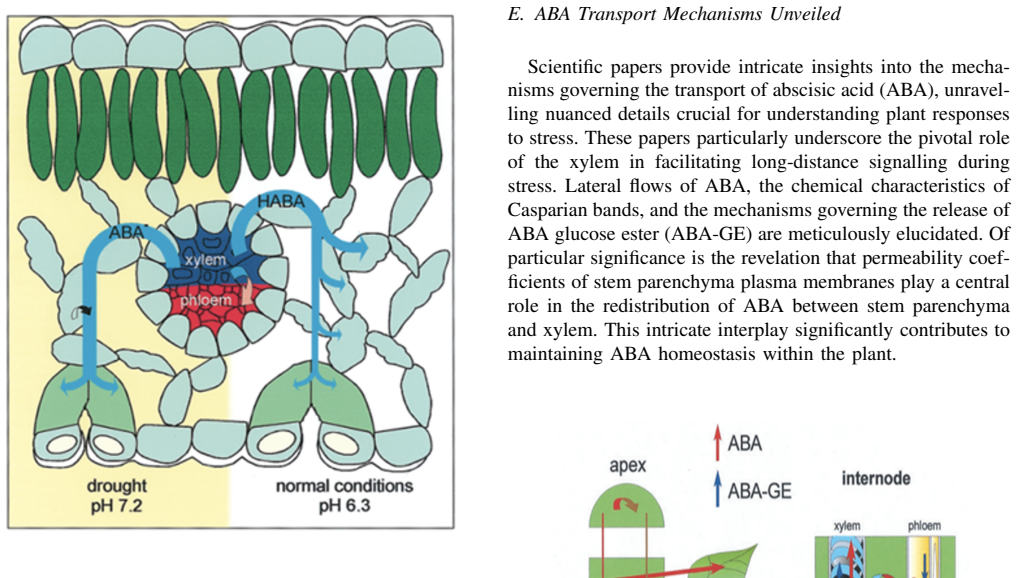
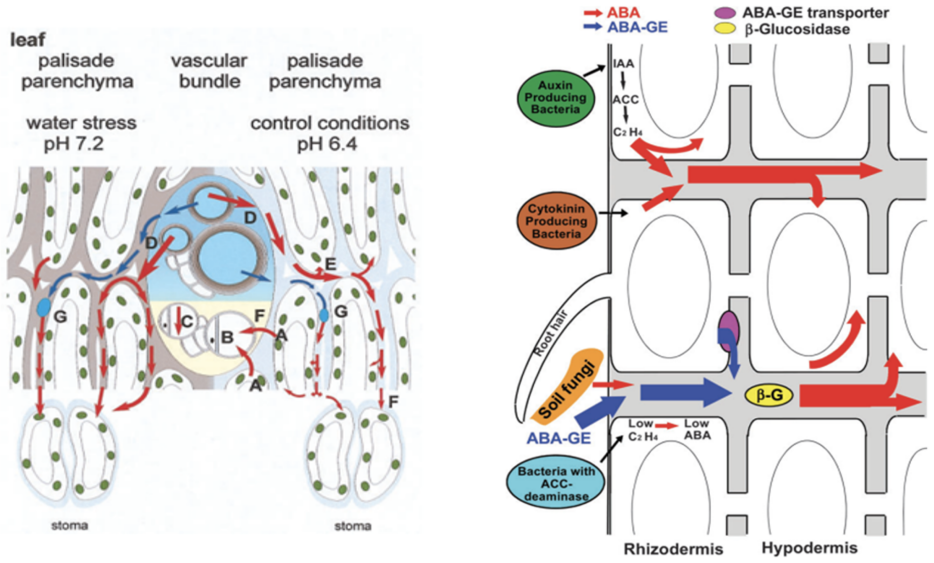
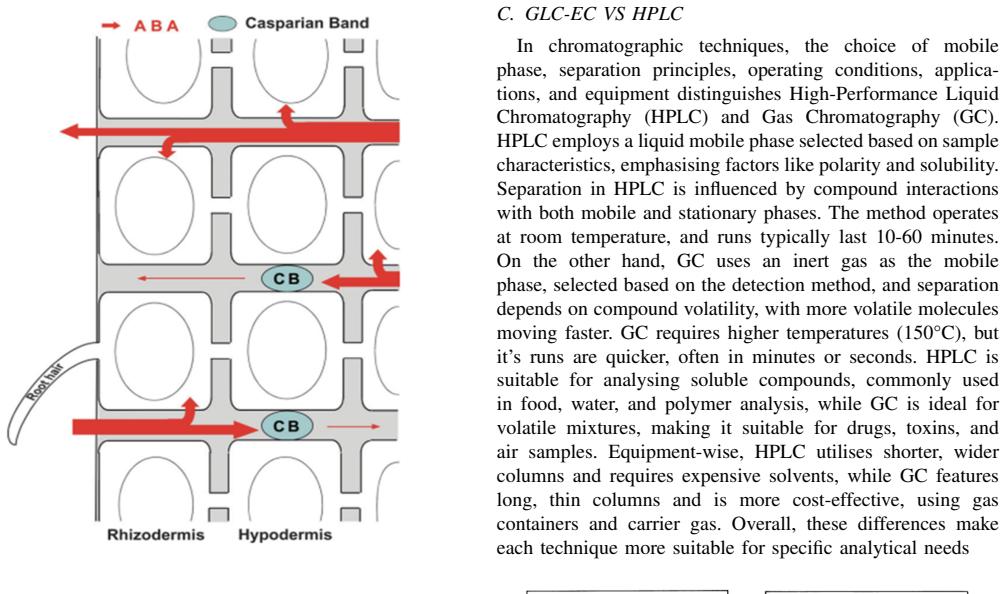
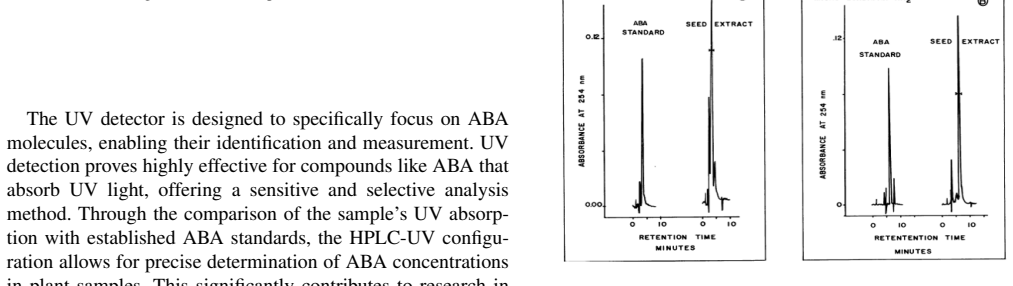
Abscisic acid (ABA) is a central plant hormone for coordinating responses to drought, salinity, cold stress, pathogen attack, wounding, and developmental aging. This paper reviews the biological stimuli that increase ABA biosynthesis, the main production sites and pathways, and the long-distance movement of ABA through plant vascular tissues. It then discusses experimental quantification approaches, including gas-liquid chromatography with electron-capture detection and high-performance liquid chromatography with ultraviolet detection. Finally, the paper presents a molecular-communication-inspired model of ABA transport in which root-side ABA release is represented as a transmitter, the xylem pathway as a bounded channel, and soybean tissue as a receiver. MATLAB Brownian-motion simulations are used to evaluate the effects of released molecule quantity and receiver radius on the detected ABA signal. The results show that higher release quantities produce smoother and stronger reception trends, while larger receivers increase molecule-capture probability.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper reviews biological stimuli for ABA biosynthesis, production sites, long-distance vascular transport, and experimental quantification methods (e.g., GLC-ECD, HPLC-UV). It then presents a molecular-communication model in which root ABA release is a transmitter, the xylem a bounded channel, and soybean tissue a receiver, followed by MATLAB Brownian-motion simulations that examine the effects of released molecule quantity and receiver radius on detected signal trends. The reported results indicate that higher release quantities yield smoother and stronger reception while larger receivers increase capture probability.
Significance. If the diffusion-only model were shown to be a reasonable proxy for xylem ABA transport, the simulation trends would provide a quantitative framework for exploring parameter effects on signaling efficiency within the molecular-communication paradigm. The interdisciplinary framing (plant biology + MC) is potentially useful for the eess.SP community, but the lack of any parameter values, error bars, experimental comparison, or advection sensitivity analysis means the work currently functions more as an exploratory modeling exercise than a validated contribution.
major comments (2)
- [Abstract (modeling paragraph)] Abstract (modeling paragraph): the central simulation results rest on a pure Brownian-motion model with a bounded channel, yet no justification is supplied for omitting advection despite xylem flow velocities of 0.1–1 mm/s and Peclet numbers ≫1; this assumption is load-bearing for any claim that the reported quantity/radius trends apply to real long-distance ABA signaling.
- [Abstract (simulation results)] Abstract (simulation results): the reported trends on release quantity and receiver radius are presented without any parameter values, error bars, statistical measures, or direct comparison to measured ABA concentrations or transport times in soybean or other species, leaving the quantitative claims unanchored to experiment.
minor comments (1)
- [Abstract] The abstract mentions specific quantification techniques (GLC-ECD, HPLC-UV) but does not indicate whether these data are used to calibrate or validate the subsequent model; a brief statement of their role would improve clarity.
Simulated Author's Rebuttal
We thank the referee for the constructive comments on our manuscript. We address each major point below, agreeing where the modeling choices require clarification and where additional reporting is needed. The work is positioned as an exploratory application of molecular communication concepts to ABA signaling, and revisions will better reflect its limitations.
read point-by-point responses
-
Referee: [Abstract (modeling paragraph)] Abstract (modeling paragraph): the central simulation results rest on a pure Brownian-motion model with a bounded channel, yet no justification is supplied for omitting advection despite xylem flow velocities of 0.1–1 mm/s and Peclet numbers ≫1; this assumption is load-bearing for any claim that the reported quantity/radius trends apply to real long-distance ABA signaling.
Authors: We acknowledge that advection dominates xylem transport and that the pure Brownian-motion model is a significant simplification. The manuscript frames the work within the molecular-communication paradigm and intentionally isolates diffusive effects in a bounded channel for initial exploration. No claim is made that the trends apply directly to in-vivo conditions. In revision we will add explicit justification for the modeling choice, state that the reported quantity and radius effects are specific to the diffusion-only case, and note that advection-diffusion extensions remain future work. This addresses the load-bearing nature of the assumption without altering the core simulations. revision: partial
-
Referee: [Abstract (simulation results)] Abstract (simulation results): the reported trends on release quantity and receiver radius are presented without any parameter values, error bars, statistical measures, or direct comparison to measured ABA concentrations or transport times in soybean or other species, leaving the quantitative claims unanchored to experiment.
Authors: We agree that parameter values and variability measures should be reported. The revised manuscript will list the diffusion coefficient, channel dimensions, time-step size, molecule counts, and receiver radii used, together with the number of independent Brownian-motion runs performed. Mean reception curves will be accompanied by standard-deviation bands. Because the study is a modeling exercise, direct numerical comparison to measured ABA concentrations or transport times is not provided; literature values for ABA levels will be cited to contextualize the chosen parameters, but new experimental anchoring lies outside the present scope. revision: partial
- Direct quantitative comparison of simulated signals to measured ABA concentrations and transport times in soybean, which would require new experimental data collection beyond the modeling focus of the manuscript.
Circularity Check
No circularity: simulation outputs are independent of inputs
full rationale
The paper's central results are numerical outputs from MATLAB Brownian-motion simulations that directly vary release quantity and receiver radius as independent parameters and record the resulting reception trends and capture probabilities. These quantities are not fitted to data and then re-predicted; they are simulation inputs whose effects are measured. No equations reduce a claimed prediction to the input by construction, no self-citations are load-bearing for the modeling choices, and no uniqueness theorems or ansatzes are imported. The derivation chain is therefore self-contained: model definition followed by forward simulation.
Axiom & Free-Parameter Ledger
free parameters (2)
- released molecule quantity
- receiver radius
axioms (2)
- domain assumption Molecule transport in the xylem can be approximated by Brownian motion inside a bounded channel.
- domain assumption Soybean tissue functions as a passive receiver that detects arriving ABA molecules.
Reference graph
Works this paper leans on
-
[1]
Jasmonoyl isoleucine accumu- lation is needed for abscisic acid build- up in roots of Arabidopsis under water stress conditions
Ollas C, Arbona V , G ´oMez-Cadenas A. Jasmonoyl isoleucine accumu- lation is needed for abscisic acid build- up in roots of Arabidopsis under water stress conditions. Plant Cell Environ 2015;38:2157–2170
2015
-
[2]
Embryophyte stress signaling evolved in the algal progenitors of land plants
Vries J, Curtis BA, Gould SB, Archibald JM. Embryophyte stress signaling evolved in the algal progenitors of land plants. Proc Natl Acad Sci USA 2018;115:E3471–E3480
2018
-
[3]
Gibberellin requirement for Arabidopsis seed germination is determined both by testa characteristics and em- bryonic abscisic acid
Debeaujon I, Koornneef M. Gibberellin requirement for Arabidopsis seed germination is determined both by testa characteristics and em- bryonic abscisic acid. Plant Physiol 2000;122:415–424
2000
-
[4]
The Arabidopsis DELAY OF GERMI- NATION 1 gene affects ABSCISIC ACID INSENSITIVE 5 (ABI5) expression and genetically interacts with ABI3 during Arabidopsis seed development
Dekkers BJ, He H, Hanson J, Willems LA, Jamar DC, Cueff G, Rajjou L, Hilhorst HW, Bentsink L. The Arabidopsis DELAY OF GERMI- NATION 1 gene affects ABSCISIC ACID INSENSITIVE 5 (ABI5) expression and genetically interacts with ABI3 during Arabidopsis seed development. Plant J 2016;85:451–465
2016
-
[5]
Regulation of seed dormancy by abscisic acid and DELAY OF GERMINATION 1
Dekkers BJW, Bentsink L. Regulation of seed dormancy by abscisic acid and DELAY OF GERMINATION 1. Seed Sci Res 2015;25:82–98
2015
-
[6]
Seed germination of GA-insensitive sleepy1 mutants does not require RGL2 protein disappearance in Arabidopsis
Ariizumi T, Steber CM. Seed germination of GA-insensitive sleepy1 mutants does not require RGL2 protein disappearance in Arabidopsis. Plant Cell 2007;19:791–804
2007
-
[7]
Guard cell sensory systems: Recent insights on stomatal responses to light, abscisic acid, and CO2
Assmann SM, Jegla T. Guard cell sensory systems: Recent insights on stomatal responses to light, abscisic acid, and CO2. Curr Opin Plant Biol 2016;33:157–167
2016
-
[8]
Localisation and expression of zeaxanthin epoxidase mRNA in Arabidopsis in response to drought stress and during seed development
Audran C, Liotenberg S, Gonneau M, North H, Frey A, Tap-Waksman K, Vartanian N, Marion-Poll A. Localisation and expression of zeaxanthin epoxidase mRNA in Arabidopsis in response to drought stress and during seed development. Funct Plant Biol 2001;28:1161
2001
-
[9]
In vitro reconstitution of an abscisic acid signalling pathway
Fujii H, Chinnusamy V , Rodrigues A, Rubio S, Antoni R, Park SY , Cutler SR, Sheen J, Rodriguez PL, Zhu JK. In vitro reconstitution of an abscisic acid signalling pathway. Nature 2009;462:660–664
2009
-
[10]
Arabidopsis mutant deficient in 3 ab- scisic acid- activated protein kinases reveals critical roles in growth, reproduction, and stress
Fujii H, Zhu JK. Arabidopsis mutant deficient in 3 ab- scisic acid- activated protein kinases reveals critical roles in growth, reproduction, and stress. Proc Natl Acad Sci USA 2009;106:8380–8385
2009
-
[11]
Abscisic Acid
Nambara E, Brian T, Murray BG, Murphy DJ. Abscisic Acid. In: Thomas B, Murray BG, Murphy DJ, editors. Encyclopedia of Applied Plant Sciences (Second Edition). Academic Press; 2017. p. 361–366. ISBN: 9780123948083
2017
-
[12]
Abscisic Acid as Pathogen Effector and Immune Regulator
Lievens L, Pollier J, Goossens A. Abscisic Acid as Pathogen Effector and Immune Regulator. [No further information available]
-
[13]
Long-distance signalling of abscisic acid (ABA): the factors regulating the intensity of the ABA signal
Jiang F, Hartung W. Long-distance signalling of abscisic acid (ABA): the factors regulating the intensity of the ABA signal. Journal of Experimental Botany. 2008;59(1):37–43
2008
-
[14]
Abscisic acid in the xylem: where does it come from, where does it go to? Journal of Experimental Botany
Hartung W, Sauter A, Hose E. Abscisic acid in the xylem: where does it come from, where does it go to? Journal of Experimental Botany. 2002;53(366):27–32
2002
-
[15]
Early-stopped aggregation: Adaptive inference with computational efficiency
Nambara E, Marion-Poll A. Abscisic Acid Biosynthesis And Catabolism. Annu Rev Plant Biol. 2005;56:165-85. Doi: 10.1146/An- nurev.Arplant.56.032604.144046. Pmid: 15862093
work page internal anchor Pith review Pith/arXiv arXiv doi:10.1146/an- 2005
-
[16]
Abscisic Acid: Biosynthesis, Inactivation, Homoeostasis And Signalling
Dong T, Park Y , Hwang I. Abscisic Acid: Biosynthesis, Inactivation, Homoeostasis And Signalling. Essays Biochem. 2015;58:29-48. Doi: 10.1042/Bse0580029. Pmid: 26374885
-
[17]
The Arabidopsis Aba-Deficient Mutant Aba4 Demon- strates That The Major Route For Stress-Induced Aba Accumulation Is Via Neoxanthin Isomers
North Hm, De Almeida A, Boutin Jp, Frey A, To A, Botran Sotta B, Marion-Poll A. The Arabidopsis Aba-Deficient Mutant Aba4 Demon- strates That The Major Route For Stress-Induced Aba Accumulation Is Via Neoxanthin Isomers. Plant J 2007;50:810–824
2007
-
[18]
A Unique Short-Chain Dehydrogenase/Reductase In Arabidopsis Glucose Signaling And Ab- scisic Acid Biosynthesis And Functions
Cheng Wh, Endo A, Zhou L, Penney J, Chen Hc, Arroyo A, Leon P, Nambara E, Asami T, Seo M, Koshiba T, Sheen J. A Unique Short-Chain Dehydrogenase/Reductase In Arabidopsis Glucose Signaling And Ab- scisic Acid Biosynthesis And Functions. Plant Cell 2002;14:2723–2743
2002
-
[19]
AREB1 is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerance in Arabidopsis
Fujita Y , Fujita M, Satoh R, Maruyama K, Parvez MM, Seki M, Hiratsu K, Ohme-Takagi M, Shinozaki K, Yamaguchi-Shinozaki K. AREB1 is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerance in Arabidopsis. Plant Cell 2005;17:3470–3488
2005
-
[20]
Archetypal roles of an abscisic acid receptor in drought and sugar responses in liverworts
Jahan A, Komatsu K, Wakida-Sekiya M, Hiraide M, Tanaka K, Ohtake R, Umezawa T, Toriyama T, Shinozawa A, Yotsui I, Sakata Y , Takezawa D. Archetypal roles of an abscisic acid receptor in drought and sugar responses in liverworts. Plant Physiol 2019;179:317–328
2019
-
[21]
Overexpression of PYL5 in rice enhances drought tolerance, inhibits growth, and modulates gene expression
Kim H, Lee K, Hwang H, Bhatnagar N, Kim DY , Yoon IS, Byun MO, Kim ST, Jung KH, Kim BG. Overexpression of PYL5 in rice enhances drought tolerance, inhibits growth, and modulates gene expression. J Exp Bot 2014;65:453–464
2014
-
[22]
Phytohormone mutants in plant research
Reid JB. Phytohormone mutants in plant research. J Plant Growth Regul 1990;9:97
1990
-
[23]
CLE25 peptide regulates phloem initiation in Arabidopsis through a CLERK-CLV2 receptor complex
Ren SC, Song XF, Chen WQ, Lu R, Lucas WJ, Liu CM. CLE25 peptide regulates phloem initiation in Arabidopsis through a CLERK-CLV2 receptor complex. J Integr Plant Biol 2019;61:1043–1061
2019
-
[24]
The absence of histone H2B monoubiq- uitination in the Arabidopsis hub1 (rdo4) mutant reveals a role for chromatin remodeling in seed dormancy
Liu Y , Koornneef M, Soppe WJ. The absence of histone H2B monoubiq- uitination in the Arabidopsis hub1 (rdo4) mutant reveals a role for chromatin remodeling in seed dormancy. Plant Cell 2007;19:433–444
2007
-
[25]
UDP-glucosyltransferase71c5, a major glucosyltrans- ferase, mediates abscisic acid homeostasis in Arabidopsis
Liu Z, Yan JP, Li DK, Luo Q, Yan Q, Liu ZB, Ye LM, Wang JM, Li XF, Yang Y . UDP-glucosyltransferase71c5, a major glucosyltrans- ferase, mediates abscisic acid homeostasis in Arabidopsis. Plant Physiol 2015;167:1659–1670
2015
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.