Predator-associated cues promote host riding, and coupling to mobile hosts improves survival in an epizoic limpet
Pith reviewed 2026-06-26 06:04 UTC · model grok-4.3
The pith
Crab cues increase limpet attachment to hosts while riding mobile hosts raises survival rates.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
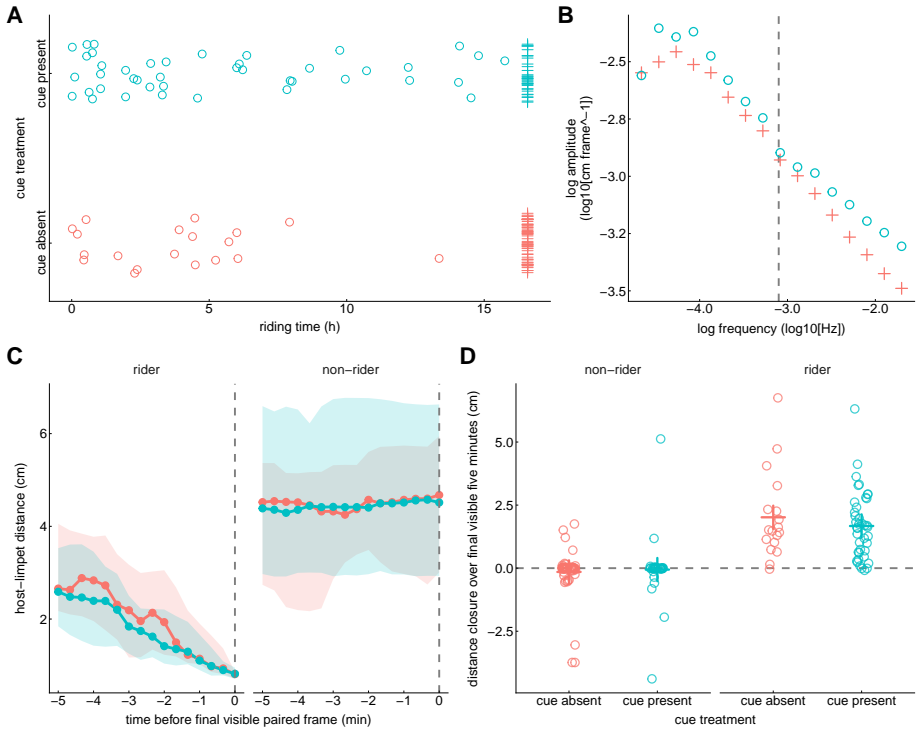
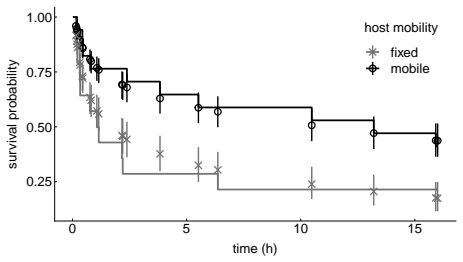
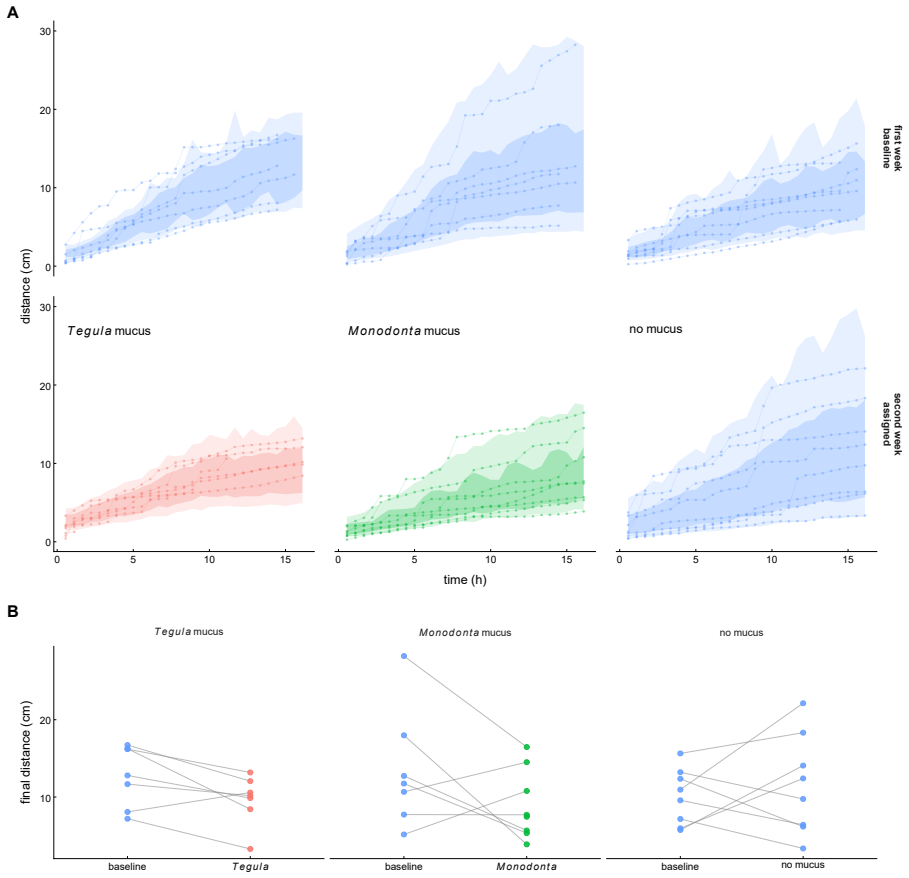
In the host-riding assay crab-associated cues raised attachment counts and shallowed the high-frequency tail of the locomotor amplitude spectrum (posterior median slope from -0.353 to -0.287). Paired-trajectory analysis showed stronger distance closure under the same cues. The survival assay produced a posterior median hazard ratio of 2.111 for fixed versus mobile hosts and posterior median survival probabilities of 0.437 on mobile hosts versus 0.175 on fixed hosts. Mucus-conditioned surfaces reduced the final cumulative-distance range of single limpets relative to controls.
What carries the argument
Host-riding response to predator cues, quantified by attachment counts, spectral analysis of locomotion, visible paired-trajectory distance closure, and survival hazard models.
If this is right
- Crab cues raise attachment rates inside the observation window.
- Paired trajectories show a measurable pre-riding approach component that strengthens under cues.
- Attachment to mobile hosts produces lower mortality than attachment to fixed hosts.
- Host mucus alone can restrict the final distance range traveled by solitary limpets.
Where Pith is reading between the lines
- Limpets appear able to use chemical predator signals to decide when to initiate host-seeking movement.
- The same cue-driven attachment logic could operate in other epizoic marine invertebrates facing mobile predators.
- Direct field counts of attachment frequency across gradients of crab density would test whether the lab pattern scales to natural populations.
Load-bearing premise
The cue and host-mobility treatments isolate crab signals and mobility without unmeasured differences in water chemistry, host identity, or limpet condition altering attachment or survival.
What would settle it
Repeating the host-riding assay and finding no difference in attachment rates between cue-present and cue-absent chambers would falsify the claim that predator cues promote riding.
Figures
read the original abstract
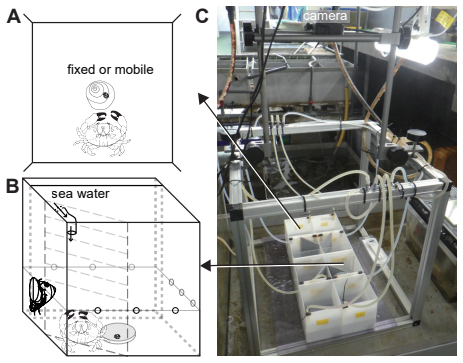
Epizoic limpets may reduce predation risk by riding mobile gastropod hosts, but the behavioral steps leading to host attachment and the benefits of attachment remain poorly understood. We examined these issues in the epizoic limpet $\textit{Lottia tenuisculpta}$ using a host-riding assay, paired-trajectory analyses of pre-riding movement, a survival assay, and mucus-conditioning assays. In the host-riding experiment, involving 156 limpets in 39 chambers, crab-associated cues increased attachment within the observation window from 19 of 80 individuals in the cue-absent treatment to 42 of 76 individuals in the cue-present treatment. In the same assay, the high-frequency tail of the locomotor amplitude spectrum became shallower under cue-present conditions, with the posterior median slope shifting from -0.353 to -0.287. Direct analysis of visible paired host-limpet trajectories further showed stronger distance closure under crab-associated cues. Distance closure was quantified over the final visible five minutes before riding or before the final visible paired frame. In the survival assay, based on 31 valid trials, the fitted model indicated lower survival after attachment on fixed hosts than on mobile hosts: the posterior median hazard ratio for fixed versus mobile hosts was 2.111, and posterior median survival at the end of the observation window was 0.437 on mobile hosts but 0.175 on fixed hosts. In a separate single-limpet locomotion assay, gastropod mucus-conditioned surfaces yielded narrower final cumulative-distance ranges than the no-mucus control. Together, these results indicate that predator-associated cues promote host riding, visible paired trajectories reveal a pre-riding approach component, coupling to mobile hosts improves survival, and host-associated surface cues may narrow solitary-limpet movement.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper examines behavioral and survival benefits of host riding in the epizoic limpet Lottia tenuisculpta. Using a host-riding assay with 156 limpets, it reports that crab-associated cues raise attachment from 19/80 to 42/76 individuals and shallow the high-frequency tail of the locomotor amplitude spectrum (posterior median slope from -0.353 to -0.287). Paired-trajectory analysis shows stronger distance closure under cues. A survival assay with 31 valid trials yields a posterior median hazard ratio of 2.111 for fixed vs mobile hosts, with survival probabilities 0.437 (mobile) vs 0.175 (fixed). Mucus-conditioning assays indicate narrower cumulative-distance ranges on conditioned surfaces.
Significance. If the central results hold, the work supplies direct experimental counts and posterior estimates linking predator cues to increased host attachment, pre-riding approach behavior, and a survival advantage for mobile-host coupling in an epizoic system. The multi-assay design and concrete sample sizes (156 limpets, 31 trials) are strengths that allow falsifiable claims about behavioral ecology. The survival finding, however, rests on a small sample whose robustness is not fully documented in the provided text.
major comments (1)
- [Abstract, survival assay description] Abstract, survival assay paragraph: the claim that coupling to mobile hosts improves survival rests on a model fitted to only 31 valid trials that produces a posterior median hazard ratio of 2.111 and survival probabilities 0.437 vs 0.175. No event counts, credible intervals, censoring details, or sensitivity checks to baseline hazard or priors are supplied; in survival analysis this sample size is small enough that a few influential observations or model choices could alter the result, making the estimate load-bearing for the survival-benefit conclusion.
minor comments (2)
- [Abstract] Abstract: the phrasing 'the high-frequency tail of the locomotor amplitude spectrum became shallower' and the slope values (-0.353 to -0.287) would benefit from a brief definition of the amplitude spectrum or a reference to the methods section where it is defined.
- [Abstract] Abstract: the mucus-conditioning result is stated only qualitatively ('narrower final cumulative-distance ranges'); adding the actual ranges or a statistical summary would improve clarity.
Simulated Author's Rebuttal
We thank the referee for their thoughtful review and for highlighting the need for greater transparency in the survival assay. We address the single major comment below and will incorporate additional details and checks in a revised manuscript.
read point-by-point responses
-
Referee: [Abstract, survival assay description] Abstract, survival assay paragraph: the claim that coupling to mobile hosts improves survival rests on a model fitted to only 31 valid trials that produces a posterior median hazard ratio of 2.111 and survival probabilities 0.437 vs 0.175. No event counts, credible intervals, censoring details, or sensitivity checks to baseline hazard or priors are supplied; in survival analysis this sample size is small enough that a few influential observations or model choices could alter the result, making the estimate load-bearing for the survival-benefit conclusion.
Authors: We agree that the survival assay relies on a modest number of valid trials (31) and that the submitted manuscript does not report event counts, credible intervals, censoring information, or sensitivity analyses. These omissions make it difficult for readers to evaluate robustness. In the revision we will add: (i) the observed number of events (deaths) in the fixed-host and mobile-host groups, (ii) 95% credible intervals for the hazard ratio and for the reported survival probabilities, (iii) explicit description of any right-censoring that occurred, and (iv) results of sensitivity checks that vary the baseline hazard specification and the prior distributions. These additions will be placed in both the Methods and Results sections so that the survival-benefit claim can be assessed more fully while preserving the original experimental design and sample size. revision: yes
Circularity Check
No circularity: results from direct experiments and standard statistical analysis of new data
full rationale
The paper presents empirical findings from new assays (host-riding with 156 limpets, survival with 31 trials, mucus-conditioning, paired trajectories). Attachment rates, spectral slopes, distance closure, and hazard ratios are measured or fitted directly to the collected data. No equations, self-citations, or prior parameters are invoked such that any reported outcome reduces to a quantity defined by construction from the inputs. The derivation chain is self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
free parameters (1)
- posterior median hazard ratio
axioms (1)
- standard math Bayesian survival model assumptions hold for the hazard-ratio estimation
Reference graph
Works this paper leans on
-
[1]
Stan: A probabilistic programming language.Journal of Statistical Software76:1–32. URL: https: //doi.org/10.18637/jss.v076.i01 Chialvo DR
-
[2]
Emergent complex neural dynamics.Nature Physics6:744–750. URL:https://doi.org/10. 1038/nphys1803 ClinchyM,SheriffMJ,ZanetteLY.2013. Predator-inducedstressandtheecologyoffear.FunctionalEcology27:56–65. URL:https://doi.org/10.1111/1365-2435.12007 Codling EA, Plank MJ, Benhamou S
-
[3]
URL:https://doi.org/10.1098/rsif.2008.0014 Dalesman S, Rundle SD, Coleman RA, Cotton PA
Random walk models in biology.Journal of the Royal Society Interface 5:813–834. URL:https://doi.org/10.1098/rsif.2008.0014 Dalesman S, Rundle SD, Coleman RA, Cotton PA
-
[4]
URL: https://doi.org/10.1016/j.anbehav.2005
Cue association and antipredator behaviour in a pulmonate snail,Lymnaeastagnalis.AnimalBehaviour71:789–797. URL: https://doi.org/10.1016/j.anbehav.2005. 05.028 Dalesman S, Rundle SD, Cotton PA
-
[5]
URL:https://doi.org/10.1111/ j.1365-2427.2007.01843.x Gelman A, Hill J
Predator regime influences innate anti-predator behaviour in the freshwater gastropodLymnaea stagnalis.Freshwater Biology52:2134–2140. URL:https://doi.org/10.1111/ j.1365-2427.2007.01843.x Gelman A, Hill J. 2007.Data Analysis Using Regression and Multilevel/Hierarchical Models. Cambridge University Press. URL:https://doi.org/10.1017/CBO9780511790942 Houck...
-
[6]
URL: https://doi.org/10.1111/j.0030-1299.2004.12369.x Kagaya K, Nakano T, Nakayama R
Antipredator behaviour mediated by chemical cues: the role of conspecific alarm signalling and predator labelling in the avoidance response of a marine gastropod.Oikos104:43–50. URL: https://doi.org/10.1111/j.0030-1299.2004.12369.x Kagaya K, Nakano T, Nakayama R
-
[7]
URL:https://doi.org/10.20965/ jrm.2025.p0099 Kano Y
Multiple power laws and scaling relation in exploratory locomotion of the snailTegula nigerrima.Journal of Robotics and Mechatronics37:99–104. URL:https://doi.org/10.20965/ jrm.2025.p0099 Kano Y
2025
-
[8]
URL:https://doi.org/10.1098/rsbl.2009.0191 Kats LB, Dill LM
Hitchhiking behaviour in the obligatory upstream migration of amphidromous snails.Biology Letters 5:465–468. URL:https://doi.org/10.1098/rsbl.2009.0191 Kats LB, Dill LM
-
[9]
URL: https://doi.org/10.1080/11956860.1998.11682468 Lima SL
Chemosensory assessment of predation risk by prey animals.Écoscience5:361–394. URL: https://doi.org/10.1080/11956860.1998.11682468 Lima SL
-
[10]
URL: https://doi.org/10.2307/1313225 Lima SL, Dill LM
Nonlethal effects in the ecology of predator-prey interactions.BioScience48:25–34. URL: https://doi.org/10.2307/1313225 Lima SL, Dill LM
-
[11]
URL:https://doi.org/10.1139/z90-092 Mapstone BD, Underwood AJ, Creese RG
Behavioral decisions made under the risk of predation: a review and prospectus.Canadian Journal of Zoology68:619–640. URL:https://doi.org/10.1139/z90-092 Mapstone BD, Underwood AJ, Creese RG
-
[12]
Experimental analyses of the commensal relation between intertidal gastropodsPatelloida mufriaand the trochidAustrocochlea constricta.Marine Ecology Progress Series17:85–100. URL:https://doi.org/10.3354/meps017085 Mathis A, Mamidanna P, Cury KM, Abe T, Murthy VN, Mathis MW, Bethge M
-
[13]
DeepLabCut: markerless pose estimation of user-defined body parts with deep learning.Nature Neuroscience21:1281–1289. URL: https://doi.org/10.1038/s41593-018-0209-y Morales JM, Haydon DT, Frair J, Holsinger KE, Fryxell JM
-
[14]
URL:https://doi.org/10.1890/ 03-0269 NakayamaR,NakanoT,AsakuraA.2022
Extracting more out of relocation data: building movement models as mixtures of random walks.Ecology85:2436–2445. URL:https://doi.org/10.1890/ 03-0269 NakayamaR,NakanoT,AsakuraA.2022. SubstratevarietyandhostpreferenceoftheepizoiclimpetLottiatenuisculpta (Patellogastropoda: Lottiidae).Molluscan Research42:31–40. URL:https://doi.org/10.1080/13235818. 2022.2...
-
[15]
URL:https://doi.org/10.18941/venus.83.1-4_111 10 Nakayama R, Nakano T, Yusa Y
Selective settlement of the planktonic larvae of the epizoic limpetLottia tenuisculpta (Patellogastropoda: Lottiidae).Venus83:111–120. URL:https://doi.org/10.18941/venus.83.1-4_111 10 Nakayama R, Nakano T, Yusa Y
-
[16]
Seasonal utilization patterns of two snail hosts by the epizoic limpet Lottia tenuisculpta(Gastropoda: Patellogastropoda).Journal of Experimental Marine Biology and Ecology 530–531:151402. URL:https://doi.org/10.1016/j.jembe.2020.151402 Nathan R, Getz WM, Revilla E, Holyoak M, Kadmon R, Saltz D, Smouse PE
-
[17]
A movement ecology paradigm for unifying organismal movement research.Proceedings of the National Academy of Sciences of the United States of America105:19052–19059. URL:https://doi.org/10.1073/pnas.0800375105 Nicholson-JackAE,HarrisJL,BallardK,TurnerKME,StevensGMW.2021. Ahitchhikerguidetomantarays: patterns of association betweenMobula alfredi,M. birostr...
-
[18]
URL: https://doi.org/10.1016/j.tree.2007.10.009 Preisser EL, Bolnick DI, Benard MF
State-space models of individual animal movement.TrendsinEcology&Evolution23:87–94. URL: https://doi.org/10.1016/j.tree.2007.10.009 Preisser EL, Bolnick DI, Benard MF
-
[19]
URL:https://doi.org/10.1890/04-0719 Singer JD, Willett JB
Scared to death? The effects of intimidation and consumption in predator-prey interactions.Ecology86:501–509. URL:https://doi.org/10.1890/04-0719 Singer JD, Willett JB
-
[20]
URL: https://doi.org/10.1111/j.1461-0248.2004.00719.x Stan Development Team
Local adaptation in host use among marine invertebrates.Ecology Letters8:448–459. URL: https://doi.org/10.1111/j.1461-0248.2004.00719.x Stan Development Team. 2026.Stan Reference Manual, Version 2.38. URL: https://mc-stan.org/docs/ reference-manual/ Turner AM, Bernot RJ, Boes CM
-
[21]
URL:https://doi.org/10.1034/j.1600-0706
Chemical cues modify species interactions: the ecological consequences of predator avoidance by freshwater snails.Oikos88:148–158. URL:https://doi.org/10.1034/j.1600-0706. 2000.880117.x Turner AM, Fetterolf SA, Bernot RJ
-
[22]
URL:https://www.biodiversitylibrary
Good hosts and their guests: relations between trochid gastropods and the epizoic limpetCrepidula adunca.The Nautilus101:69–74. URL:https://www.biodiversitylibrary. org/page/8097562 Wada Y, Noda T, Ida TY, Iwatani Y, Sato T
-
[23]
Tracing the battle: role of mucus trails in information warfare between predator snail and prey limpet.Journal of Animal Ecology95:851–864. URL:https://doi.org/10. 1111/1365-2656.70235 Wahl M
-
[24]
Marine epibiosis. I. Fouling and antifouling: some basic aspects.Marine Ecology Progress Series 58:175–189. URL:https://doi.org/10.3354/meps058175 Wahl M
-
[25]
URL:https://doi.org/10.1080/08927010802339772 Werner EE, Peacor SD
Ecological lever and interface ecology: epibiosis modulates the interactions between host and environment.Biofouling24:427–438. URL:https://doi.org/10.1080/08927010802339772 Werner EE, Peacor SD
-
[26]
URL:https://doi.org/10.1890/0012-9658(2003)084%5B1083:AROTII%5D2.0.CO;2 Yarnall JL
A review of trait-mediated indirect interactions in ecological communities.Ecology 84:1083–1100. URL:https://doi.org/10.1890/0012-9658(2003)084%5B1083:AROTII%5D2.0.CO;2 Yarnall JL
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.