Modelling the evolution of traits in a two-sex population, with an application to grandmothering
Pith reviewed 2026-05-25 10:04 UTC · model grok-4.3
The pith
Grandmothering alone can evolve longer lifespans for both sexes via a two-sex fitness landscape.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
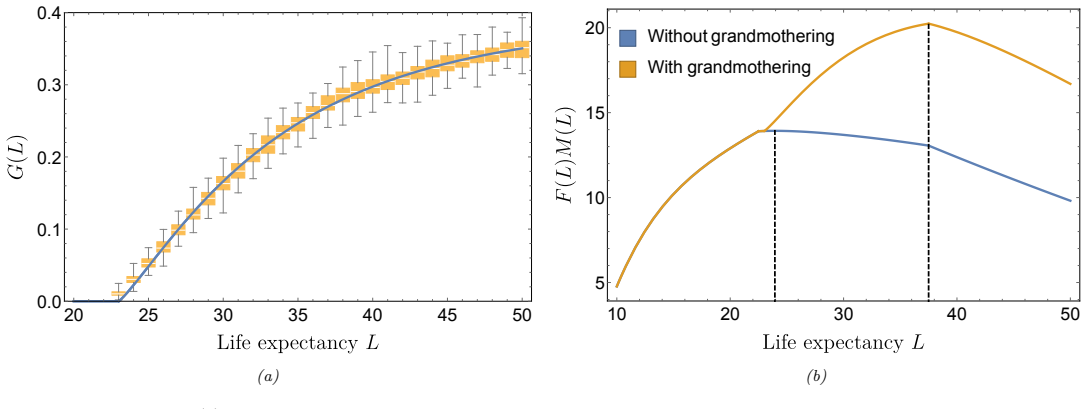
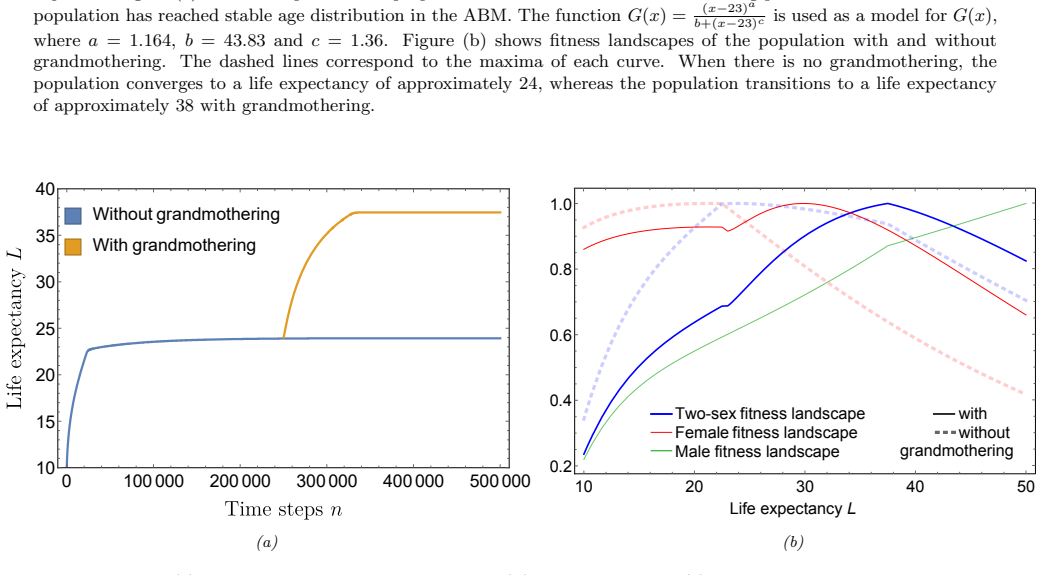
The paper derives a formula for the fitness landscape of a heritable trait in a two-sex population and shows that grandmothering, by lengthening female post-fertile longevity and raising sex ratios in fertile ages, produces higher net fitness for both males and females at extended lifespans, thereby supplying an evolutionary trajectory to greater longevities.
What carries the argument
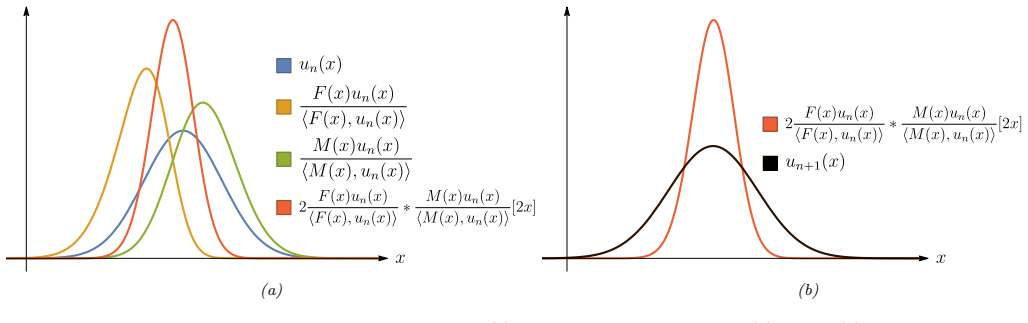
The fitness landscape formula obtained by treating trait evolution as a dynamical system that generalizes the Price Equation to two sexes and age structure.
If this is right
- Grandmothering shifts the population toward higher overall sex ratios through extended female post-fertile life.
- Males gain a longer fertile span as longevity increases.
- Both sexes experience a net fitness increase at longer lifespans.
- Grandmothering supplies a self-contained evolutionary route to higher longevities without requiring additional selective pressures.
Where Pith is reading between the lines
- The same landscape formula could be used to compare grandmothering against other forms of kin help in different species.
- Predictions about age-specific sex ratios offer a measurable signature that could be checked against demographic records from societies with varying grandmother involvement.
- The model implies that once grandmothering appears, selection can favor further increases in longevity even if other life-history changes remain fixed.
Load-bearing premise
Post-fertile females' provisioning of grandchildren must lengthen female post-reproductive longevity in a manner that raises net fitness for both sexes.
What would settle it
Empirical data showing that grandmother provisioning fails to increase the reproductive success of daughters or their offspring enough to favor longer female post-reproductive life would refute the claimed trajectory.
Figures
read the original abstract
We present a mathematical simplification for the evolutionary dynamics of a heritable trait within a two-sex population. This trait is assumed to control the timing of sex-specific life-history events, such as the age of sexual maturity and end of female fertility, and each sex has a distinct fitness tradeoff associated with the trait. We provide a formula for the fitness landscape of the population and show a natural extension of the result to an arbitrary number of heritable traits. Our method can be viewed as a dynamical systems generalisation of the Price Equation to include two sexes{, age structure} and multiple traits. We use this formula to examine the effect of grandmothering, whereby post-fertile females subsidize their daughter's fertility by provisioning grandchildren. Grandmothering can drive a shift towards higher sex ratios due to lengthening female post-fertile longevity, leading to changes in fitness for both sexes. For males, increased longevity is accompanied by a substantially longer fertile lifespan resulting in higher sex ratios in the fertile ages. Our fitness landscapes show a net increase in fitness for both males and females at longer lifespans, and as a result, we find that grandmothering alone provides an evolutionary trajectory to higher longevities.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper derives a formula for the fitness landscape of a heritable trait controlling sex-specific life-history events (e.g., age at maturity and end of female fertility) in a two-sex population. It presents this as a dynamical-systems generalization of the Price equation that incorporates two sexes, age structure, and multiple traits, and applies the formula to grandmothering, where post-reproductive females provision grandchildren. The central claim is that grandmothering produces a net fitness increase for both sexes at longer post-fertile lifespans, thereby supplying an evolutionary trajectory toward higher longevities and altered sex ratios.
Significance. If the derivation is internally consistent and the grandmothering subsidy emerges from the trait dynamics rather than being inserted as an independent parameter, the work supplies a compact analytic tool for two-sex age-structured evolution that could be applied to other life-history questions. The explicit extension to multiple traits and the reported fitness-landscape results for grandmothering would constitute a concrete, falsifiable prediction about the direction of selection on longevity.
major comments (2)
- [Application section] Application section (grandmothering model): the subsidy is introduced as an additive benefit to daughter's fertility that lengthens post-fertile longevity. It is not shown whether this term is a derived function of the heritable trait that sets the end of female fertility or an externally parameterized quantity. If the latter, the reported net fitness gain at longer lifespans is an input to the landscape rather than an output of the Price-style dynamics, which directly affects the claim that grandmothering alone supplies the evolutionary trajectory.
- [Main derivation / fitness-landscape formula] Fitness-landscape formula (main derivation): the abstract states that the landscapes 'show' a net increase for both sexes, yet the provided description does not indicate how the two-sex, age-structured extension propagates the subsidy through the trait-controlled fertility schedule without additional free parameters. Clarification is required on whether the increase survives when the subsidy strength is tied to the same heritable trait that governs end-of-fertility age.
minor comments (1)
- [Abstract / multi-trait extension] The abstract mentions an extension to an arbitrary number of heritable traits but does not illustrate the multi-trait case with a concrete example or matrix notation; adding one would improve readability.
Simulated Author's Rebuttal
We thank the referee for their careful reading and constructive comments on the modeling choices in the grandmothering application. We address each major comment below with clarifications on how the subsidy enters the dynamics.
read point-by-point responses
-
Referee: [Application section] Application section (grandmothering model): the subsidy is introduced as an additive benefit to daughter's fertility that lengthens post-fertile longevity. It is not shown whether this term is a derived function of the heritable trait that sets the end of female fertility or an externally parameterized quantity. If the latter, the reported net fitness gain at longer lifespans is an input to the landscape rather than an output of the Price-style dynamics, which directly affects the claim that grandmothering alone supplies the evolutionary trajectory.
Authors: The heritable trait directly sets the age at end of female fertility and therefore the length of the post-fertile interval. The subsidy enters as an additive term to daughter fertility that is active only while the grandmother remains alive and post-fertile; its total integrated effect on the next generation is therefore a function of the trait value. The fitness landscape is obtained by substituting the resulting age-specific vital rates into the two-sex dynamical system and computing the selection gradient; the net gain at longer post-fertile lifespans is consequently an output of those dynamics rather than an exogenous input. We will add an explicit sentence in the application section stating that the subsidy duration is trait-controlled. revision: partial
-
Referee: [Main derivation / fitness-landscape formula] Fitness-landscape formula (main derivation): the abstract states that the landscapes 'show' a net increase for both sexes, yet the provided description does not indicate how the two-sex, age-structured extension propagates the subsidy through the trait-controlled fertility schedule without additional free parameters. Clarification is required on whether the increase survives when the subsidy strength is tied to the same heritable trait that governs end-of-fertility age.
Authors: The two-sex, age-structured extension inserts the grandmothering term into the fertility schedule of reproductive females; the magnitude of that term at each age is proportional to the number of post-fertile grandmothers, whose abundance is governed by the same trait that sets the end of fertility. No extra free parameters are required beyond the fixed per-year provisioning rate. The reported net fitness increase for both sexes is obtained directly from this construction. We acknowledge that making the provisioning rate itself a function of post-fertile duration would constitute a stronger test; the present model keeps the rate constant and still yields the directional result. We will insert a short paragraph after the landscape formula clarifying the propagation and note the constant-rate assumption as a modeling choice. revision: partial
Circularity Check
No significant circularity detected
full rationale
The paper derives a two-sex age-structured generalization of the Price equation as a dynamical-systems formula for the fitness landscape of heritable traits controlling life-history timing, then applies the formula to a grandmothering model in which post-fertile females provide an explicit subsidy to daughter fertility. No equations or self-citations are exhibited that reduce the reported net fitness increase at longer lifespans to a fitted parameter, a self-defined quantity, or a load-bearing prior result by the same authors. The subsidy is an explicit modeling assumption whose consequences are computed; the evolutionary trajectory is therefore an output of the derived landscape rather than a tautology. The derivation chain remains self-contained.
Axiom & Free-Parameter Ledger
free parameters (1)
- grandmothering subsidy strength
axioms (1)
- domain assumption A heritable trait controls the timing of sex-specific life-history events and each sex experiences a distinct fitness tradeoff with that trait.
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
Our fitness landscapes show a net increase in fitness for both males and females at longer lifespans... grandmothering alone provides an evolutionary trajectory to higher longevities.
-
IndisputableMonolith/Foundation/RealityFromDistinction.leanreality_from_one_distinction unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
the birth rate b(a,L) ... is of the form B(G(L))¯b(a,L) ... m=3 so that grandmothers boost ... by a factor of 3
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
J., Crewe, P., Grafen, A., and Gratwick, R
Batty, C. J., Crewe, P., Grafen, A., and Gratwick, R. (2014). Foundations of a mathematical theory of darwinism. Journal of Mathematical Biology , 69(2):295–334
work page 2014
-
[2]
Chan, M. H., Hawkes, K., and Kim, P. S. (2016). Evolution of longevity, age at last birth and sexual conflict with grandmothering. Journal of Theoretical Biology, 393:145–157
work page 2016
-
[3]
Chapman, T., Arnqvist, G., Bangham, J., and Rowe, L. (2003). Sexual conflict. Trends in Ecology and Evolution, 18(1):41–47
work page 2003
-
[4]
Charnov, E. L. (1993). Life History Invariants: Some Explorations of Symmetry in Evolutionary Ecology . Life History Invariants: Some Explorations of Symmetry in Evolutionary Ecology. Oxford University Press
work page 1993
-
[5]
Coxworth, J. E., Kim, P. S., McQueen, J. S., and Hawkes, K. (2015). Grandmothering life histories and human pair bonding. Proceedings of the National Academy of Sciences , 112(38):11806–11811. 8
work page 2015
-
[6]
Frank, S. A. (2012). Natural selection. iv. the price equation. Journal of Evolutionary Biology , 25(6):1002– 1019
work page 2012
-
[7]
Grafen, A. (2015). Biological fitness and the Price Equation in class-structured populations. Journal of Theoretical Biology, 373:62–72
work page 2015
-
[8]
S., Kennedy, B., Bohlender, R., and Hawks, J
Hawkes, K., Kim, P. S., Kennedy, B., Bohlender, R., and Hawks, J. (2011). A reappraisal of grandmothering and natural selection. Proceedings of the Royal Society of London B: Biological Sciences, 278(1714):1936–1938
work page 2011
-
[9]
Kachel, A. F., Premo, L. S., and Hublin, J. (2010). Grandmothering and natural selection. Proceedings of the Royal Society of London B: Biological Sciences
work page 2010
-
[10]
Kim, P. S., Coxworth, J. E., and Hawkes, K. (2012). Increased longevity evolves from grandmothering. Proceedings of the Royal Society of London B: Biological Sciences
work page 2012
-
[11]
Kim, P. S., McQueen, J. S., Coxworth, J. E., and Hawkes, K. (2014). Grandmothering drives the evolution of longevity in a probabilistic model. Journal of Theoretical Biology, 353:84–94
work page 2014
-
[12]
Loo, S. L., Hawkes, K., and Kim, P. S. (2017). Evolution of male strategies with sex ratio dependent payoffs: connecting pair bonds with grandmothering. In review
work page 2017
-
[13]
Parker, G. A. (2006). Sexual conflict over mating and fertilization: an overview. Philosophical Transactions of the Royal Society B: Biological Sciences , 361(1466):235–259
work page 2006
-
[14]
Pavard, S. and Branger, F. (2012). Effect of maternal and grandmaternal care on population dynamics and human life-history evolution: a matrix projection model. Theoretical Population Biology, 82(4):364–376
work page 2012
-
[15]
Price, G. R. (1970). Selection and covariance. Nature, 227(5257):520–521
work page 1970
-
[16]
Price, G. R. (1972). Extension of covariance selection mathematics. Annals of Human Genetics, 35(4):485– 490
work page 1972
-
[17]
Schacht, R. and Bell, A. V. (2016). The evolution of monogamy in response to partner scarcity. Scientific Reports, 6:32472
work page 2016
-
[18]
S¨ uli, E. and Mayers, D. F. (2003). An Introduction to Numerical Analysis . Cambridge University Press. 5 Appendix 5.1 Stability We are interested in the equilibria of the system and how they are affected by the shape of F (x) and M(x). Specifically, we wish to find the expected value of un(x) as n→∞ . To examine this, we consider the limiting caseε→ 0, whi...
work page 2003
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.