An agent-based model of outer membrane biogenesis in Gram-negative bacteria
Pith reviewed 2026-06-28 11:57 UTC · model grok-4.3
The pith
Simulations indicate that BAM complex protein incorporation stalls and occurs only in short bursts during Gram-negative outer membrane growth.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
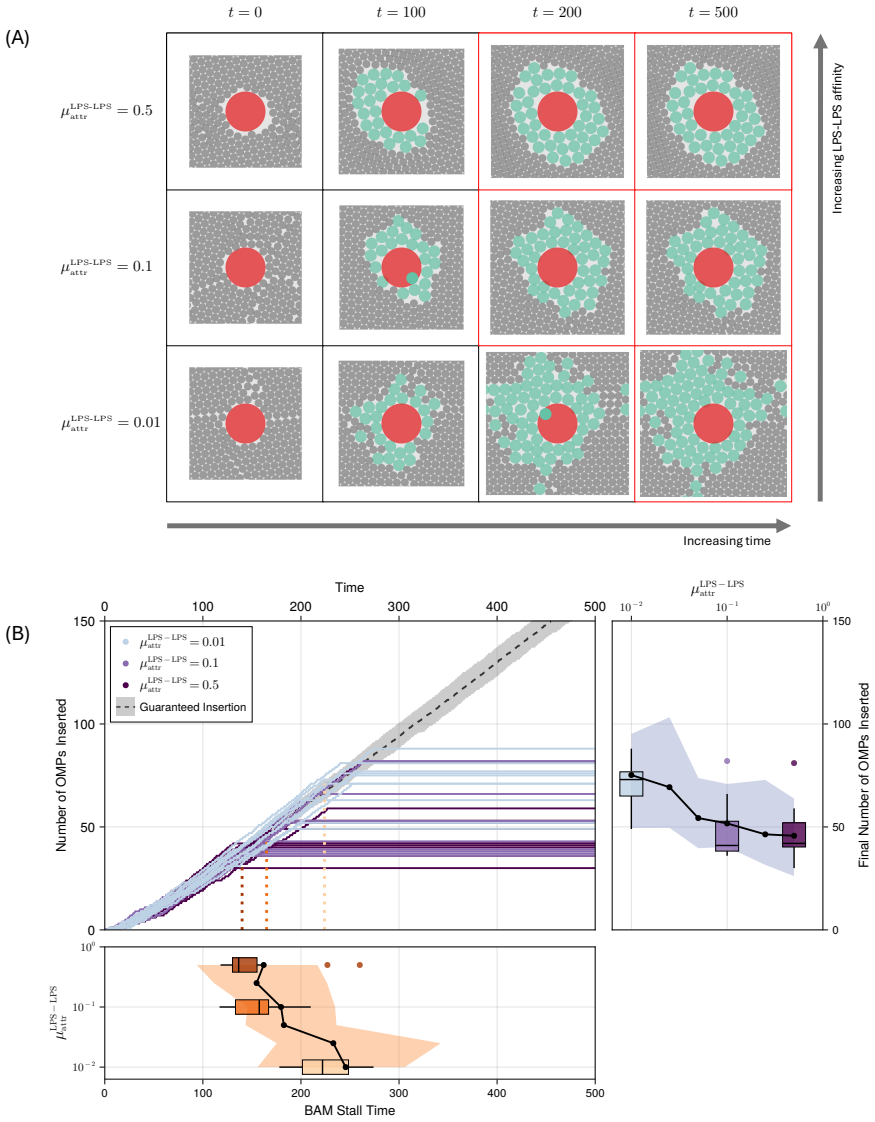
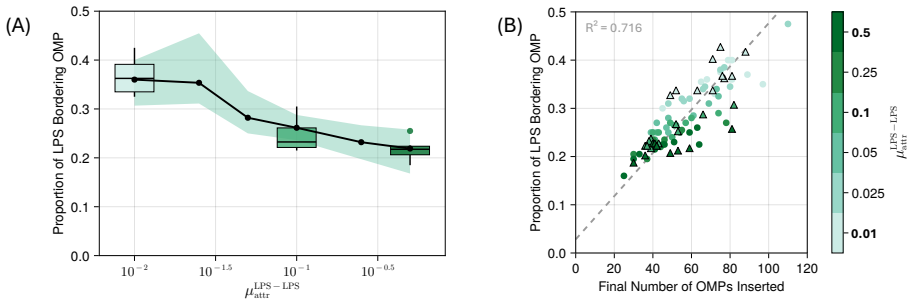
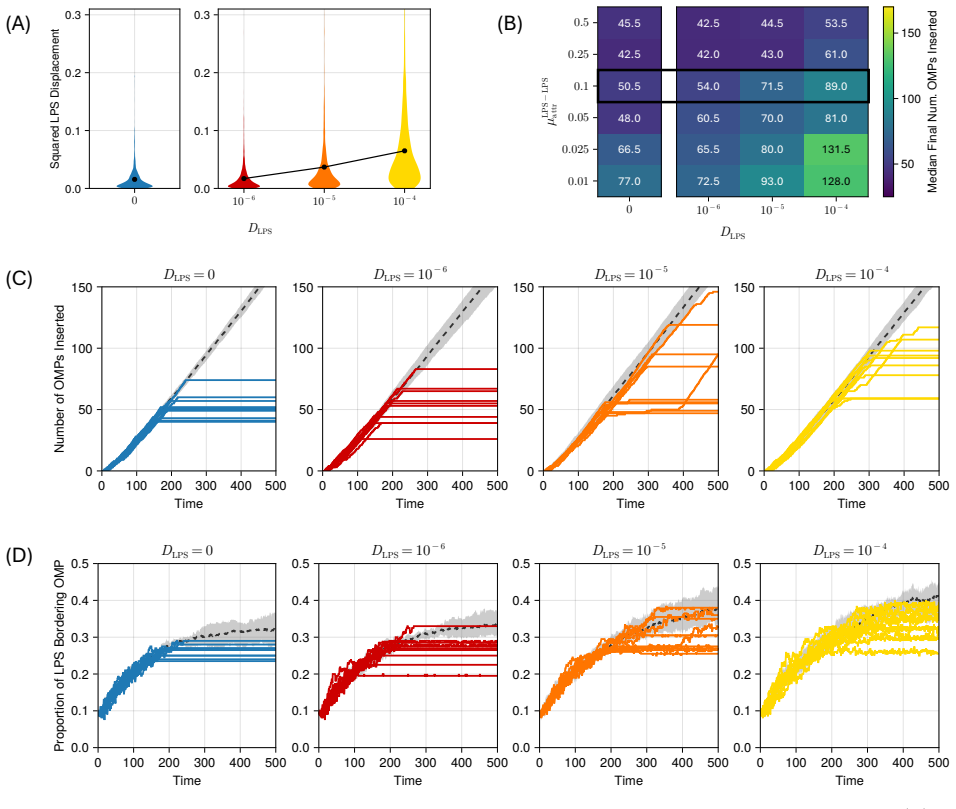
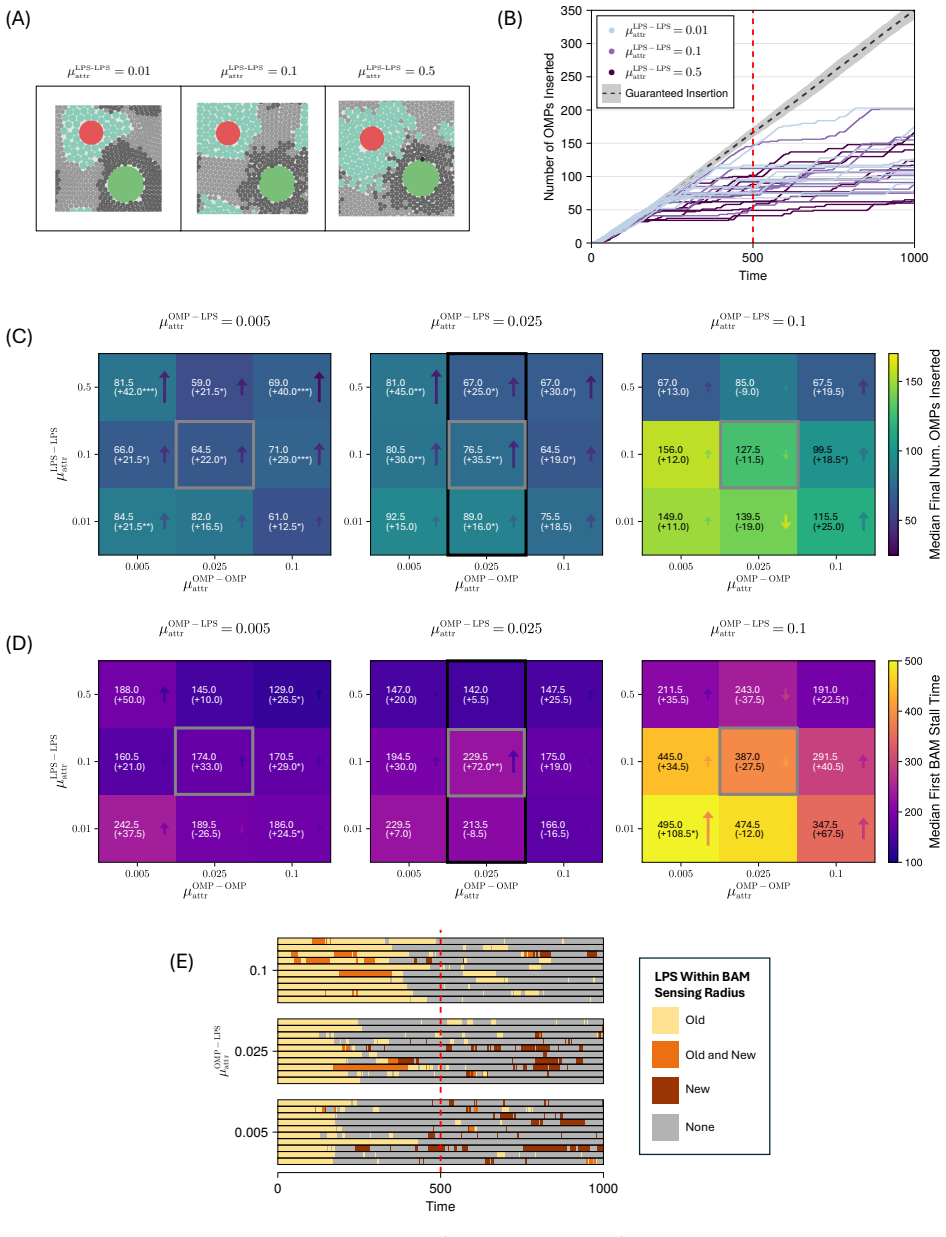
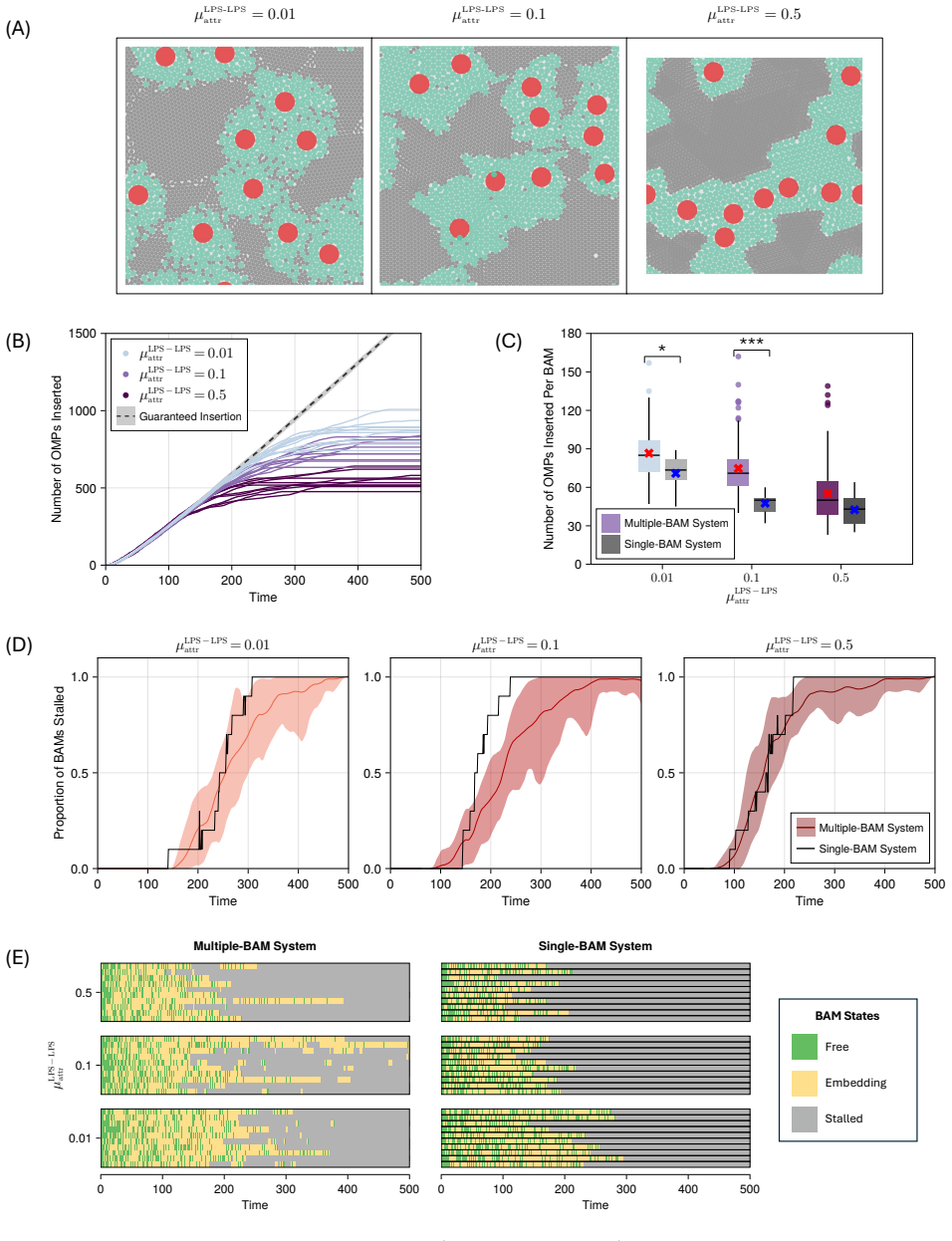
Model simulations suggest that protein incorporation into the membrane by the β-barrel assembly machinery (BAM complex) is a process which is prone to stalling, and may take place only in short bursts. We also find suggestions that BAM complexes work collaboratively with each other, and with the lipopolysaccharide-inserting Lpt complex when in close proximity.
What carries the argument
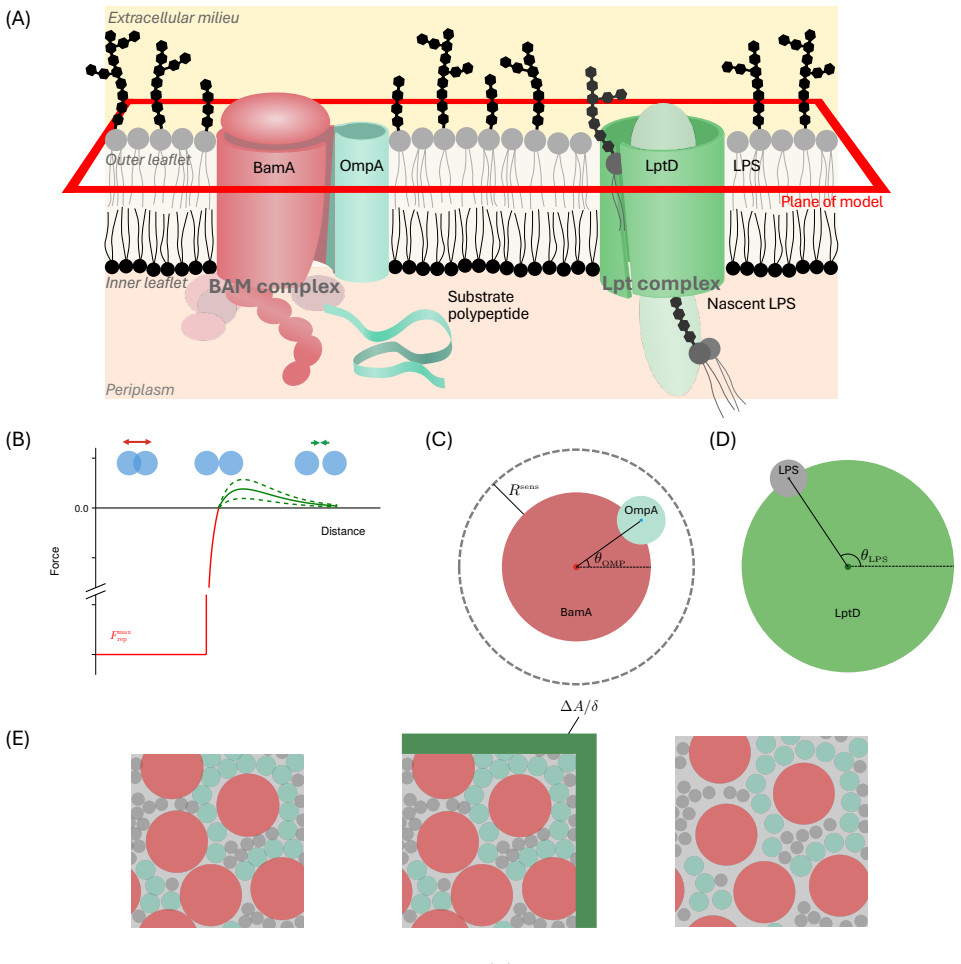
The semi-quantitative agent-based model that simulates molecular-scale dynamics of outer membrane growth over cell division timescales.
If this is right
- Protein insertion by BAM is likely intermittent rather than steady.
- BAM complexes benefit from working together in proximity.
- Coordination with Lpt complex aids efficient membrane biogenesis.
- Outer membrane growth relies on bursty rather than continuous processes.
Where Pith is reading between the lines
- If the stalling is accurate, it implies a regulatory mechanism to restart BAM activity periodically.
- The model framework could be used to predict effects of mutations or drug interventions on membrane assembly.
- Extending the model to include cell division events might reveal how membrane components are distributed to daughter cells.
Load-bearing premise
The chosen rules and parameters in the agent-based model sufficiently represent the actual dynamics of outer membrane component interactions and insertion processes.
What would settle it
Experimental data at cell-division timescales showing continuous, non-bursty protein incorporation by BAM without stalling would contradict the simulation results.
Figures
read the original abstract
The outer membrane is the interface through which Gram-negative bacteria - a broad classification of organisms including \textit{Escherichia coli} and a number of deadly pathogens - interact with the environment. Two decades of work on the process of outer membrane biogenesis have led to the discovery of the components that mediate this process, and the characterisation of structure and function of these component parts of the bacterial cell machinery. However, neither current experimental methods, nor conventional molecular dynamics (MD) simulation approaches are capable of investigating this membrane machinery on the time scale of the cell division cycle. This leaves crucial questions unanswered, such as how this lipid-poor, largely static environment is organised to permit ongoing membrane growth. Here, we introduce a semi-quantitative agent-based model to explore the molecular-scale dynamics of Gram-negative outer membrane as it grows. Model simulations across a broad region of parameter space suggest that protein incorporation into the membrane by the $\beta$-barrel assembly machinery (BAM complex) is a process which is prone to stalling, and may take place only in short bursts. We also find suggestions that BAM complexes work collaboratively with each other, and with the lipopolysaccharide-inserting Lpt complex when in close proximity. The agent-based framework we introduce provides a means to assess and generate hypotheses on outer membrane biogenesis on previously inaccessible time scales.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper introduces a semi-quantitative agent-based model to simulate outer membrane biogenesis in Gram-negative bacteria on cell-division timescales inaccessible to experiment or MD. Simulations over broad parameter space lead to the claims that BAM-mediated β-barrel insertion is prone to stalling and occurs only in short bursts, and that BAM complexes collaborate with each other and with nearby Lpt complexes.
Significance. If the model rules reasonably capture the essential molecular interactions, the work supplies a useful framework for generating hypotheses about collective membrane-assembly dynamics at long timescales. The explicit framing as exploratory hypothesis generation, rather than quantitative prediction, is a strength; the agent-based approach is well-suited to exploring stalling and proximity effects that are difficult to access otherwise.
major comments (2)
- [§3] §3 (Model Description): the stalling criterion (defined via a threshold on consecutive failed insertion attempts) is introduced without reference to any measured kinetic rates or structural data on BAM dwell times; because this definition directly generates the 'bursts' result, an explicit sensitivity analysis to the threshold value is required to establish robustness.
- [§4.2] §4.2 (Collaboration results): the reported increase in insertion rate when BAM and Lpt agents are within one interaction radius is shown only for a single radius value; the claim of collaboration therefore rests on an untested modeling choice whose variation could eliminate or reverse the effect.
minor comments (2)
- [Figure 2] Figure 2 caption should state the exact number of independent runs and the parameter ranges sampled for each panel.
- The phrase 'semi-quantitative' is used throughout but never defined operationally (e.g., which observables are matched to experiment and which are free).
Simulated Author's Rebuttal
We thank the referee for the constructive report and for recognizing the exploratory, hypothesis-generating nature of the work. We address the two major comments below with additional analyses that strengthen the robustness claims without altering the core conclusions.
read point-by-point responses
-
Referee: [§3] §3 (Model Description): the stalling criterion (defined via a threshold on consecutive failed insertion attempts) is introduced without reference to any measured kinetic rates or structural data on BAM dwell times; because this definition directly generates the 'bursts' result, an explicit sensitivity analysis to the threshold value is required to establish robustness.
Authors: We agree that the stalling threshold is a modeling choice without direct experimental calibration at present. In the revised manuscript we add an explicit sensitivity analysis (new Supplementary Figure S3) varying the consecutive-failure threshold over a factor of four around the nominal value. The short-burst insertion regime remains qualitatively intact across this range, although the precise burst length and frequency shift quantitatively; we now state this limitation and the robustness result in §3 and the discussion. revision: yes
-
Referee: [§4.2] §4.2 (Collaboration results): the reported increase in insertion rate when BAM and Lpt agents are within one interaction radius is shown only for a single radius value; the claim of collaboration therefore rests on an untested modeling choice whose variation could eliminate or reverse the effect.
Authors: The nominal interaction radius was chosen to match the approximate lateral size of the BAM and Lpt complexes (~10 nm). To test sensitivity we have now repeated the proximity analysis for radii spanning 5–20 nm (new Supplementary Figure S4). The elevation in insertion rate when a BAM and Lpt agent are co-localized persists for radii between 8 and 15 nm; outside this window the effect weakens, as expected when the radius becomes either too restrictive or too permissive. We report these bounds and the associated parameter choice explicitly in the revised §4.2. revision: yes
Circularity Check
No significant circularity; model is exploratory hypothesis generator
full rationale
The paper introduces a semi-quantitative agent-based model explicitly framed for hypothesis generation on inaccessible timescales, with claims presented as simulation suggestions rather than derivations or predictions. No equations, parameters, or self-citations are shown to reduce the central results (BAM stalling bursts, collaboration) to inputs by construction. The model rules are stated as exploratory choices, not fitted to the target conclusions, satisfying the criteria for a self-contained non-circular analysis.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
Albrecht, R., Sch \" u tz, M., Oberhettinger, P., Faulstich, M., Bermejo, I., Rudel, T., Diederichs, K., and Zeth, K. (2014). Structure of BamA, an essential factor in outer membrane protein biogenesis . Acta Crystallographica Section D , 70(6):1779--1789
2014
-
[2]
T., Poggio, S., Perry, A
Anwari, K., Webb, C. T., Poggio, S., Perry, A. J., Belousoff, M., Celik, N., Ramm, G., Lovering, A., Sockett, R. E., Smit, J., Jacobs-Wagner, C., and Lithgow, T. (2012). The evolution of new lipoprotein subunits of the bacterial outer membrane BAM complex. Molecular Microbiology , 84(5):832--844
2012
-
[3]
P., and Schirmer, T
Basl\' e , A., Rummel, G., Storici, P., Rosenbusch, J. P., and Schirmer, T. (2006). Crystal structure of osmoporin OmpC from E. coli at 2.0 A . Journal of Molecular Biology , 362(5):933--942
2006
-
[4]
Benn, G., Borrelli, C., Prakaash, D., Johnson, A. N. T., Fideli, V. A., Starr, T., Fitzmaurice, D., Combs, A. N., W \"u hr, M., Rojas, E. R., Khalid, S., Hoogenboom, B. W., and Silhavy, T. J. (2024). OmpA controls order in the outer membrane and shares the mechanical load. Proceedings of the National Academy of Sciences , 121(50):e2416426121
2024
-
[5]
Bergman, W. N. and Sousa, M. C. (2025). A fluorescent reporter and single-turnover kinetics reveal insight into BAM complex function. Proceedings of the National Academy of Sciences , 122(52):e2514687122
2025
-
[6]
J., McCarthy, J
Botos, I., Majdalani, N., Mayclin, S. J., McCarthy, J. G., Lundquist, K., Wojtowicz, D., Barnard, T. J., Gumbart, J. C., and Buchanan, S. K. (2016). Structural and functional characterization of the LPS transporter LptDE from gram-negative pathogens. Structure , 24(6):965--976
2016
-
[7]
Botos, I., Noinaj, N., and Buchanan, S. K. (2017). Insertion of proteins and lipopolysaccharide into the bacterial outer membrane. Philosophical Transactions of the Royal Society B: Biological Sciences , 372(1726):20160224
2017
-
[8]
L., Rassam, P., Birkholz, O., H \'e lie, J., Reddy, T., Beliaev, D., Hambly, B., Piehler, J., Kleanthous, C., and Sansom, M
Chavent, M., Duncan, A. L., Rassam, P., Birkholz, O., H \'e lie, J., Reddy, T., Beliaev, D., Hambly, B., Piehler, J., Kleanthous, C., and Sansom, M. S. P. (2018). How nanoscale protein interactions determine the mesoscale dynamic organisation of bacterial outer membrane proteins. Nature Communications , 9(1):2846
2018
-
[9]
L., and Sansom, M
Chavent, M., Duncan, A. L., and Sansom, M. S. (2016). Molecular dynamics simulations of membrane proteins and their interactions: from nanoscale to mesoscale. Current Opinion in Structural Biology , 40:8--16
2016
-
[10]
W., Schirmer, T., Rummel, G., Steiert, M., Ghosh, R., Pauptit, R
Cowan, S. W., Schirmer, T., Rummel, G., Steiert, M., Ghosh, R., Pauptit, R. A., Jansonius, J. N., and Rosenbusch, J. P. (1992). Crystal structures explain functional properties of two E. coli porins. Nature , 358(6389):727--733
1992
-
[11]
G., Stansfeld, P
Dong, H., Xiang, Q., Gu, Y., Wang, Z., Paterson, N. G., Stansfeld, P. J., He, C., Zhang, Y., Wang, W., and Dong, C. (2014). Structural basis for outer membrane lipopolysaccharide insertion. Nature , 511(7507):52--56
2014
-
[12]
T., Jimah, J
Doyle, M. T., Jimah, J. R., Dowdy, T., Ohlemacher, S. I., Larion, M., Hinshaw, J. E., and Bernstein, H. D. (2022). Cryo-EM structures reveal multiple stages of bacterial outer membrane protein folding. Cell , 185(7):1143--1156.e13
2022
-
[13]
Dunton, T. A. (2015). Using Molecular Simulations to Parameterize Discrete Models of Protein Movement in the Membrane . PhD thesis, University of Oxford
2015
-
[14]
Efremov, R. G. and Sazanov, L. A. (2012). Structure of Escherichia coli OmpF porin from lipidic mesophase. Journal of Structural Biology , 178(3):311--318
2012
-
[15]
B., Qiu, X., Corey, R
Fiorentino, F., Sauer, J. B., Qiu, X., Corey, R. A., Cassidy, C. K., Mynors-Wallis, B., Mehmood, S., Bolla, J. R., Stansfeld, P. J., and Robinson, C. V. (2021). Dynamics of an LPS translocon induced by substrate and an antimicrobial peptide. Nature Chemical Biology , 17(2):187--195
2021
-
[16]
H., Danoff, E
Gessmann, D., Chung, Y. H., Danoff, E. J., Plummer, A. M., Sandlin, C. W., Zaccai, N. R., and Fleming, K. G. (2014). Outer membrane -barrel protein folding is physically controlled by periplasmic lipid head groups and BamA . Proceedings of the National Academy of Sciences , 111(16):5878--5883
2014
-
[17]
D., Osorio, A., V \'a zquez-Ram \' rez, R., Arenas, T., Mendoza, L., Camarena, L., and Poggio, S
Ginez, L. D., Osorio, A., V \'a zquez-Ram \' rez, R., Arenas, T., Mendoza, L., Camarena, L., and Poggio, S. (2022). Changes in fluidity of the E. coli outer membrane in response to temperature, divalent cations and polymyxin-B show two different mechanisms of membrane fluidity adaptation. The FEBS Journal , 289(12):3550--3567
2022
-
[18]
D., Shiota, T., Stubenrauch, C
Gunasinghe, S. D., Shiota, T., Stubenrauch, C. J., Schulze, K. E., Webb, C. T., Fulcher, A. J., Dunstan, R. A., Hay, I. D., Naderer, T., Whelan, D. R., Bell, T. D. M., Elgass, K. D., Strugnell, R. A., and Lithgow, T. (2018). The WD40 protein BamB mediates coupling of BAM complexes into assembly precincts in the bacterial outer membrane. Cell Reports , 23(...
2018
-
[19]
Gutishvili, G., Yang, L., and Gumbart, J. C. (2024). Seeing is believing: Illuminating the Gram-negative outer membrane with molecular dynamics simulations. Current Opinion in Structural Biology , 87:102828
2024
-
[20]
E., Brockwell, D
Horne, J. E., Brockwell, D. J., and Radford, S. E. (2020). Role of the lipid bilayer in outer membrane protein folding in Gram-negative bacteria. Journal of Biological Chemistry , 295(30):10340--10367
2020
-
[21]
G., Schiffrin, B., White, P., Watson, M
Iadanza, M. G., Schiffrin, B., White, P., Watson, M. A., Horne, J. E., Higgins, A. J., Calabrese, A. N., Brockwell, D. J., Tuma, R., Kalli, A. C., Radford, S. E., and Ranson, N. A. (2020). Distortion of the bilayer and dynamics of the BAM complex in lipid nanodiscs. Communications Biology , 3(1):766
2020
-
[22]
and Khalid, S
Im, W. and Khalid, S. (2020). Molecular simulations of Gram-negative bacterial membranes come of age. Annual Review of Physical Chemistry , 71:171--188
2020
-
[23]
J., and Samsudin, F
Khalid, S., Piggot, T. J., and Samsudin, F. (2019). Atomistic and coarse grain simulations of the cell envelope of Gram-negative bacteria: What have we learned? Accounts of Chemical Research , 52(1):180--188
2019
-
[24]
M., Ryoo, D., Lundquist, K., and Gumbart, J
Kuo, K. M., Ryoo, D., Lundquist, K., and Gumbart, J. C. (2022). Modeling intermediates of BamA folding an outer membrane protein. Biophysical Journal , 121(17):3242--3252
2022
-
[25]
P., Clifton, L
Le Brun, A. P., Clifton, L. A., Halbert, C. E., Lin, B., Meron, M., Holden, P. J., Lakey, J. H., and Holt, S. A. (2013). Structural characterization of a model Gram-negative bacterial surface using lipopolysaccharides from rough strains of Escherichia coli. Biomacromolecules , 14(6):2014--2022
2013
-
[26]
W., Schwartz, D
Lee Upton, S., Tay, J. W., Schwartz, D. K., and Sousa, M. C. (2023). Similarly slow diffusion of BAM and SecYEG complexes in live E. coli cells observed with 3D spt-PALM . Biophysical Journal , 122(22):4382--4394
2023
-
[27]
J., Fleming, P
Lessen, H. J., Fleming, P. J., Fleming, K. G., and Sodt, A. J. (2018). Building blocks of the outer membrane: Calculating a general elastic energy model for -barrel membrane proteins. Journal of Chemical Theory and Computation , 14(8):4487--4497. PMID: 29979594
2018
-
[28]
D., Smallwood, C
Lill, Y., Jordan, L. D., Smallwood, C. R., Newton, S. M., Lill, M. A., Klebba, P. E., and Ritchie, K. (2016). Confined mobility of TonB and FepA in Escherichia coli membranes. PLOS ONE , 11(12):1--18
2016
-
[29]
J., and Stumpf, M
Lithgow, T., Stubenrauch, C. J., and Stumpf, M. P. H. (2023). Surveying membrane landscapes: a new look at the bacterial cell surface. Nature Reviews Microbiology , 21(8):502--518
2023
-
[30]
and Gumbart, J
Liu, J. and Gumbart, J. C. (2020). Membrane thinning and lateral gating are consistent features of BamA across multiple species. PLOS Computational Biology , 16(10):1--15
2020
-
[31]
F., Menzel, J., Golecki, J
M \"u hlradt, P. F., Menzel, J., Golecki, J. R., and Speth, V. (1973). Outer membrane of Salmonella. European Journal of Biochemistry , 35(3):471--481
1973
-
[32]
F., Menzel, J., Golecki, J
M \"u hlradt, P. F., Menzel, J., Golecki, J. R., and Speth, V. (1974). Lateral mobility and surface density of lipopolysaccharide in the outer membrane of Salmonella typhimurium. European Journal of Biochemistry , 43(3):533--539
1974
-
[33]
C., and Buchanan, S
Noinaj, N., Gumbart, J. C., and Buchanan, S. K. (2017). The -barrel assembly machinery in motion. Nature Reviews Microbiology , 15(4):197--204
2017
-
[34]
J., Gumbart, J
Noinaj, N., Kuszak, A. J., Gumbart, J. C., Lukacik, P., Chang, H., Easley, N. C., Lithgow, T., and Buchanan, S. K. (2013). Structural insight into the biogenesis of -barrel membrane proteins. Nature , 501(7467):385--390
2013
-
[35]
and Schulz, G
Pautsch, A. and Schulz, G. E. (2000). High-resolution structure of the OmpA membrane domain. Journal of Molecular Biology , 298(2):273--282
2000
-
[36]
H., Yang, L., Gumbart, J
Peterson, J. H., Yang, L., Gumbart, J. C., and Bernstein, H. D. (2025). Conserved lipid-facing basic residues promote the insertion of the porin OmpC into the E. coli outer membrane. mBio , 16(3):e03319--24
2025
-
[37]
A., Birkholz, O., T \'o th, C., Chavent, M., Duncan, A
Rassam, P., Copeland, N. A., Birkholz, O., T \'o th, C., Chavent, M., Duncan, A. L., Cross, S. J., Housden, N. G., Kaminska, R., Seger, U., Quinn, D. M., Garrod, T. J., Sansom, M. S. P., Piehler, J., Baumann, C. G., and Kleanthous, C. (2015). Supramolecular assemblies underpin turnover of outer membrane proteins in bacteria. Nature , 523(7560):333--336
2015
-
[38]
C., Zhang, Z., Luo, B., Xie, T., Lu, G., Zhu, X., Wei, X., Dong, C., Zhou, R., Zhang, X., Tang, X., and Dong, H
Shen, C., Chang, S., Luo, Q., Chan, K. C., Zhang, Z., Luo, B., Xie, T., Lu, G., Zhu, X., Wei, X., Dong, C., Zhou, R., Zhang, X., Tang, X., and Dong, H. (2023). Structural basis of BAM -mediated outer membrane -barrel protein assembly. Nature , 617(7959):185--193
2023
-
[39]
J., Kahne, D., and Walker, S
Silhavy, T. J., Kahne, D., and Walker, S. (2010). The bacterial cell envelope. Cold Spring Harbor Perspectives in Biology , 2(5)
2010
-
[40]
and Soares, T
Straatsma, T. and Soares, T. (2009). Characterization of the outer membrane protein OprF of Pseudomonas aeruginosa in a lipopolysaccharide membrane by computer simulation. Proteins: Structure, Function, and Bioinformatics , 74(2):475--488
2009
-
[41]
and Errington, J
Strahl, H. and Errington, J. (2017). Bacterial membranes: Structure, domains, and function. Annual Review of Microbiology , 71:519--538
2017
-
[42]
S., Trepagnier, E
Ursell, T. S., Trepagnier, E. H., Huang, K. C., and Theriot, J. A. (2012). Analysis of surface protein expression reveals the growth pattern of the Gram-negative outer membrane. PLOS Computational Biology , 8(9):1--10
2012
-
[43]
T., Heinz, E., and Lithgow, T
Webb, C. T., Heinz, E., and Lithgow, T. (2012). Evolution of the -barrel assembly machinery. Trends in Microbiology , 20(12):612--620
2012
-
[44]
N., Oluwole, A
Webby, M. N., Oluwole, A. O., Pedebos, C., Inns, P. G., Olerinyova, A., Prakaash, D., Housden, N. G., Benn, G., Sun, D., Hoogenboom, B. W., Kukura, P., Mohammed, S., Robinson, C. V., Khalid, S., and Kleanthous, C. (2022). Lipids mediate supramolecular outer membrane protein assembly in bacteria. Science Advances , 8(44):eadc9566
2022
-
[45]
W., Lundquist, K., Zhang, Z., Kuo, K
Wu, R., Bakelar, J. W., Lundquist, K., Zhang, Z., Kuo, K. M., Ryoo, D., Pang, Y. T., Sun, C., White, T., Klose, T., Jiang, W., Gumbart, J. C., and Noinaj, N. (2021). Plasticity within the barrel domain of BamA mediates a hybrid-barrel mechanism by BAM . Nature Communications , 12(1):7131
2021
-
[46]
Zahn, M., Hartmann, J.-B., and Hiller, S. (2020). Crystal structure of E. coli BamA beta-barrel with a C -terminal extension. PDB: 6FSU
2020
-
[47]
J., Green, J
Brown, P. J., Green, J. E. F., Binder, B. J., and Osborne, J. M. (2021). A rigid body framework for multicellular modeling. Nature Computational Science , 1(11):754--766
2021
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.