Evolution of longevity, age at last birth and sexual conflict with grandmothering
Pith reviewed 2026-05-25 13:51 UTC · model grok-4.3
The pith
A two-sex model finds grandmothering enables the shift from great-ape to hunter-gatherer longevities via two stable equilibria without extending the end of fertility.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
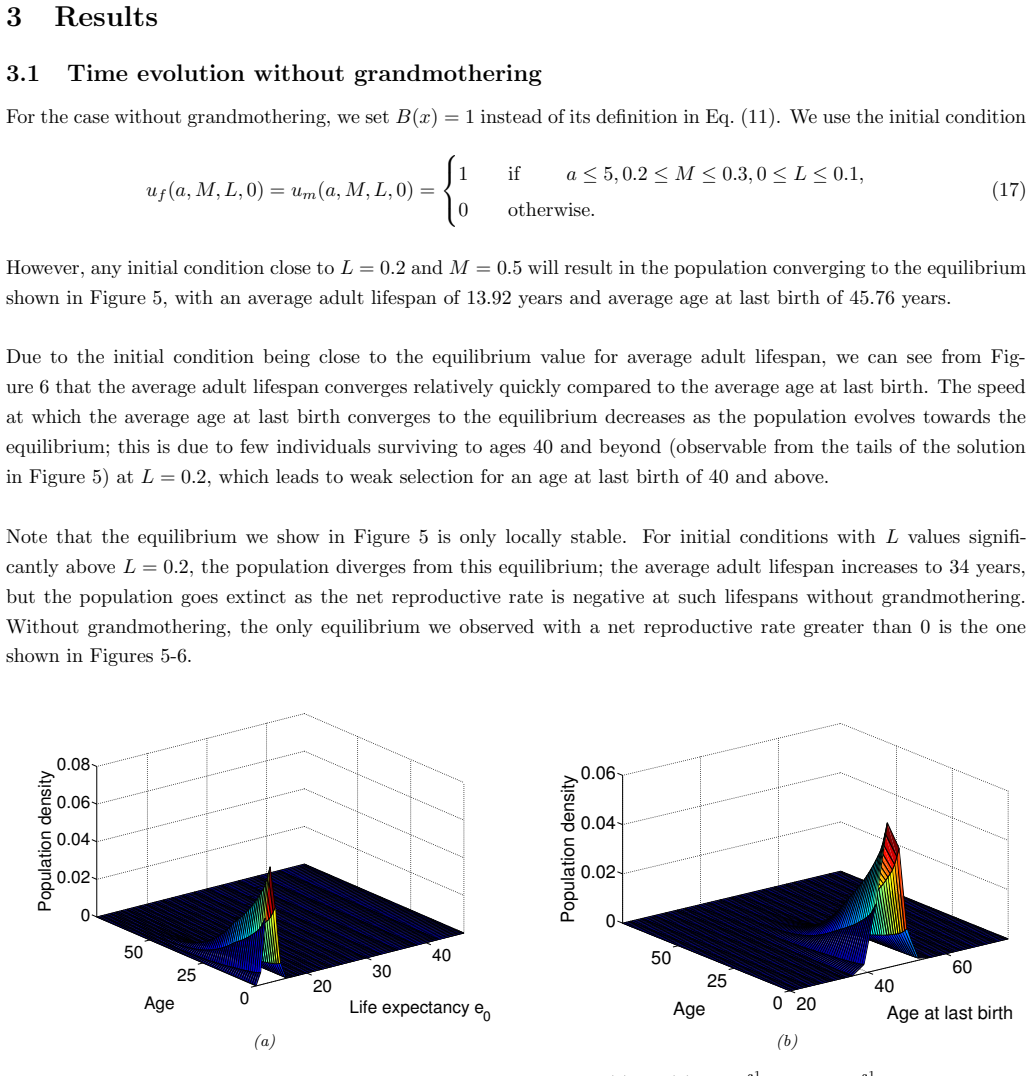
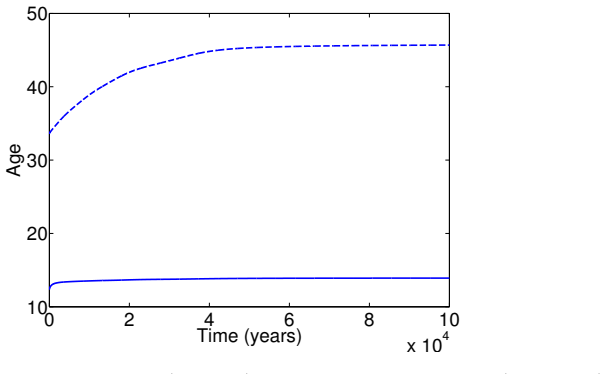
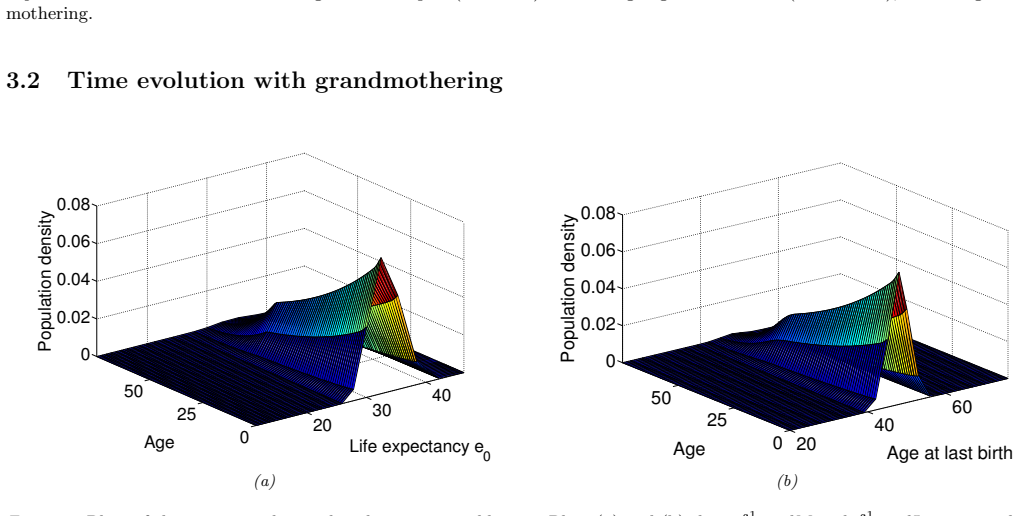
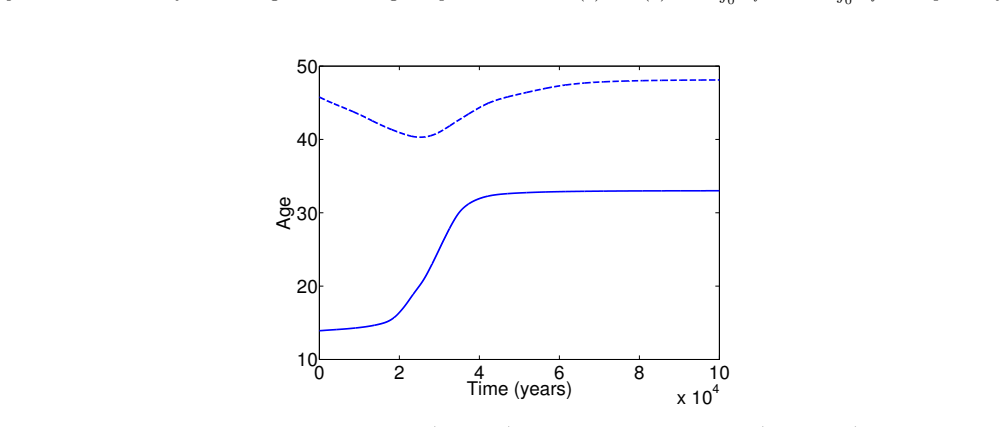
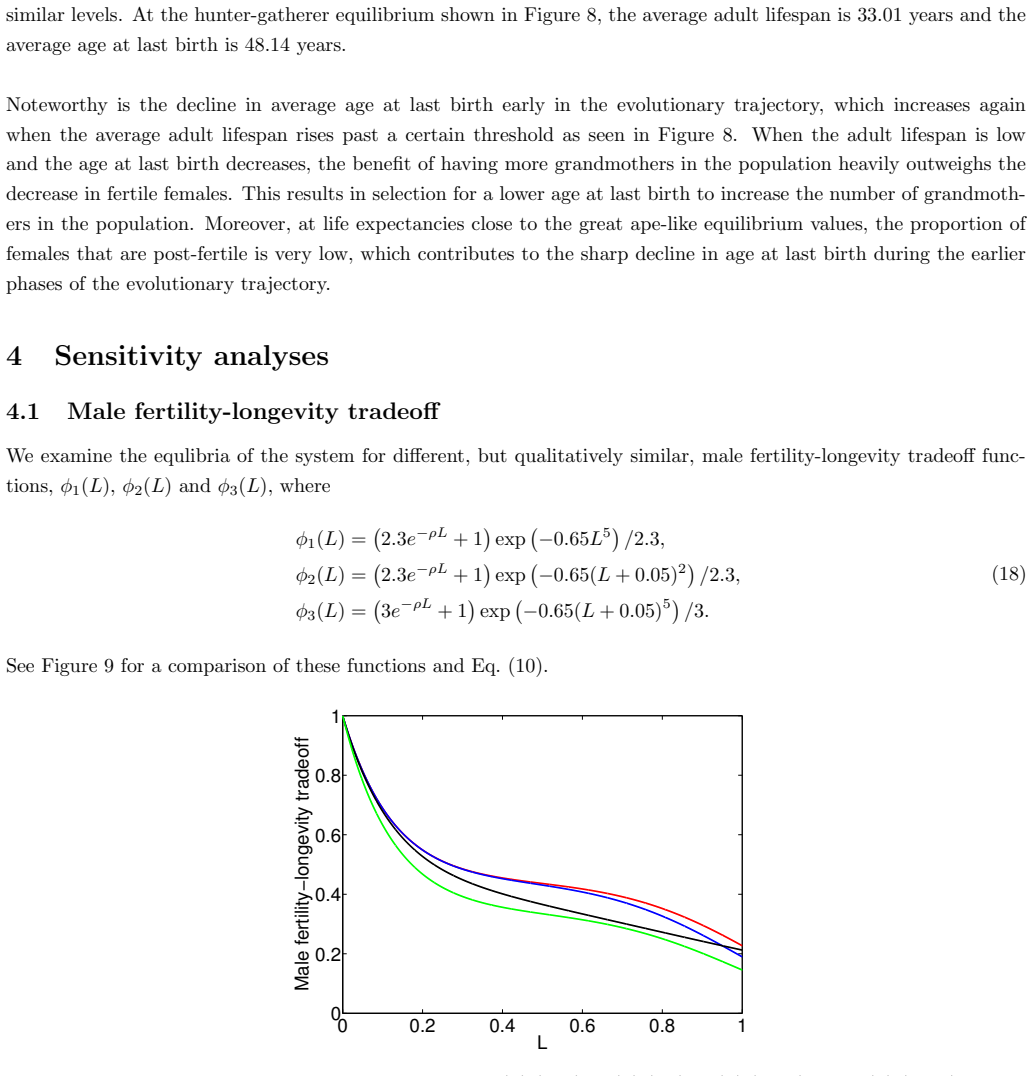
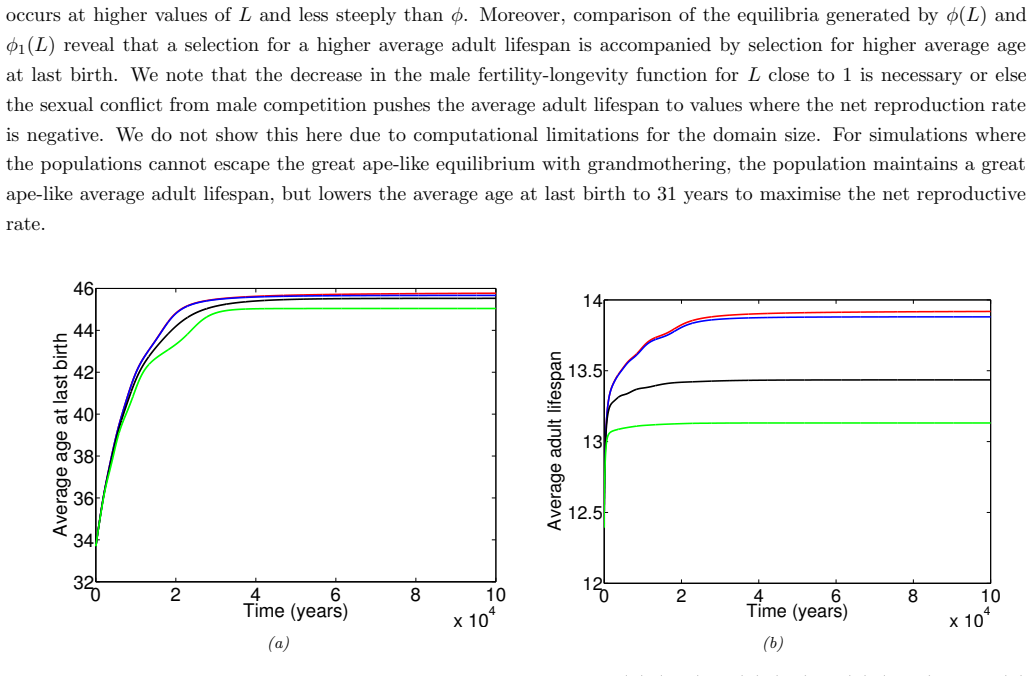
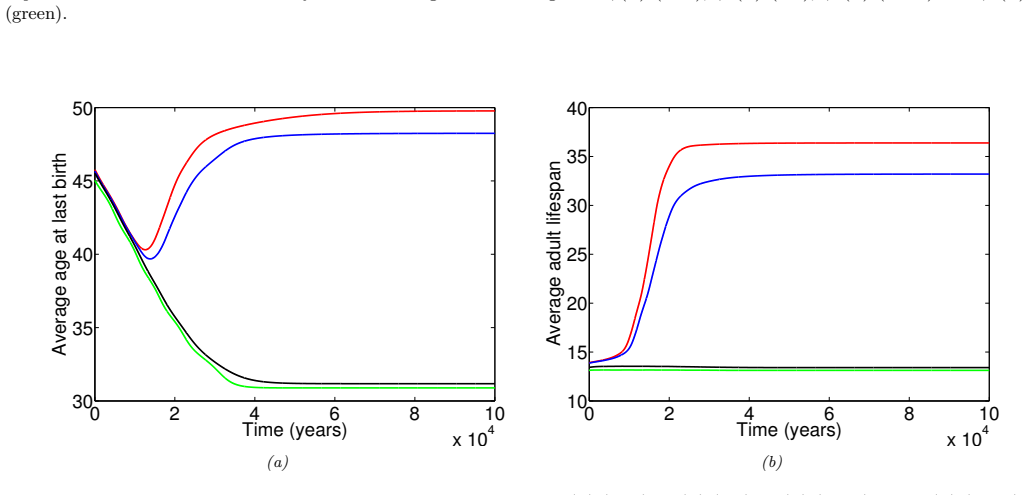
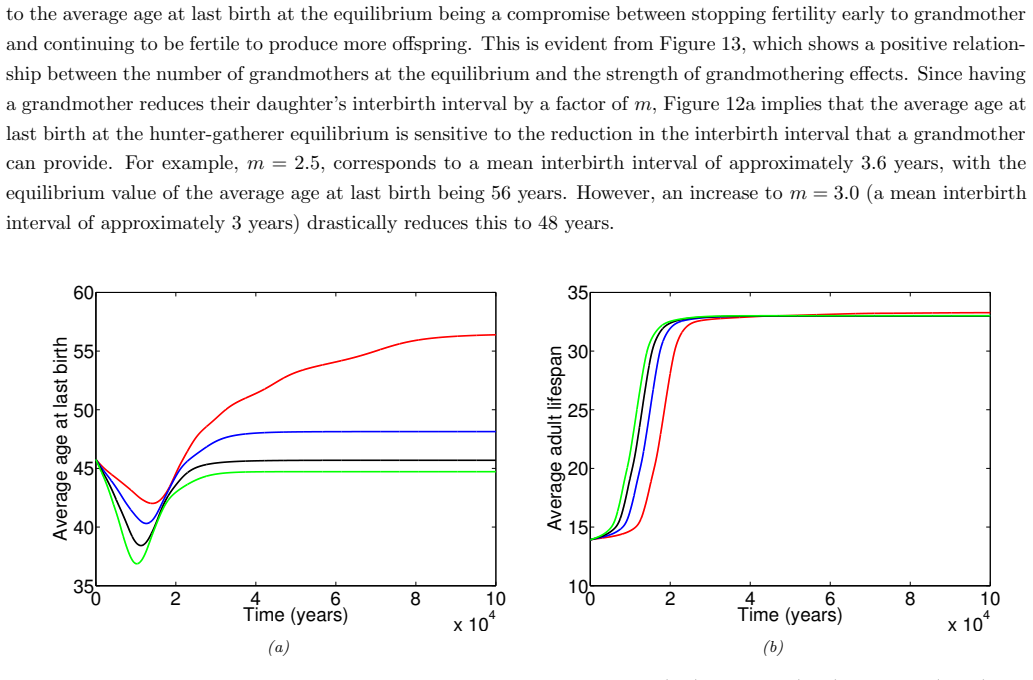
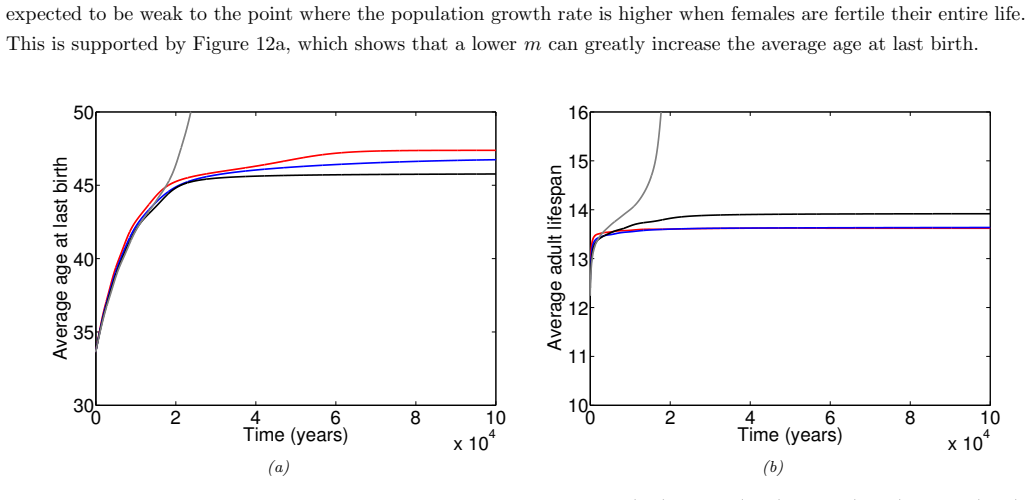
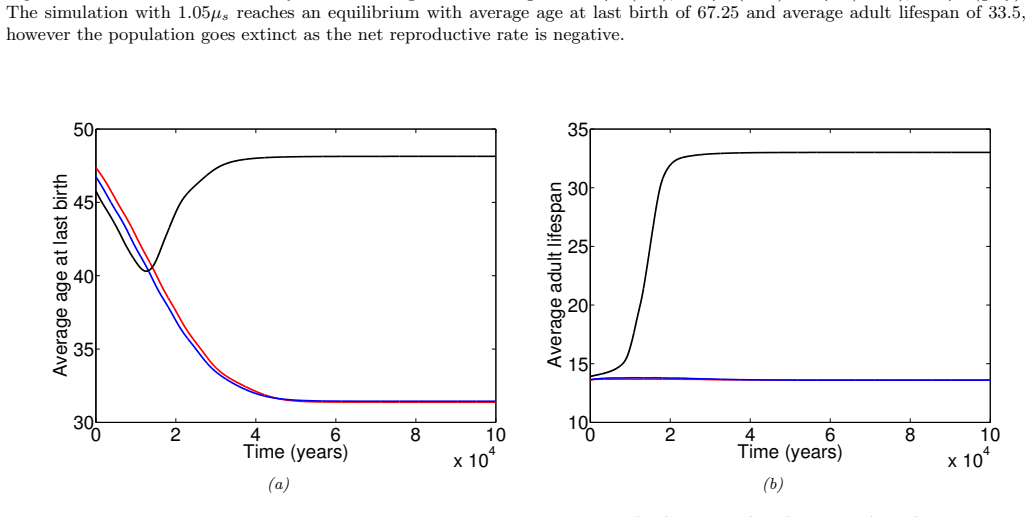
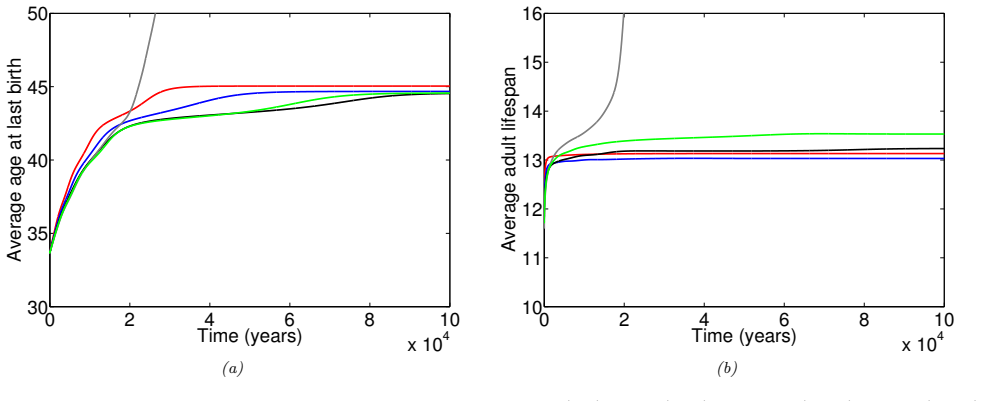
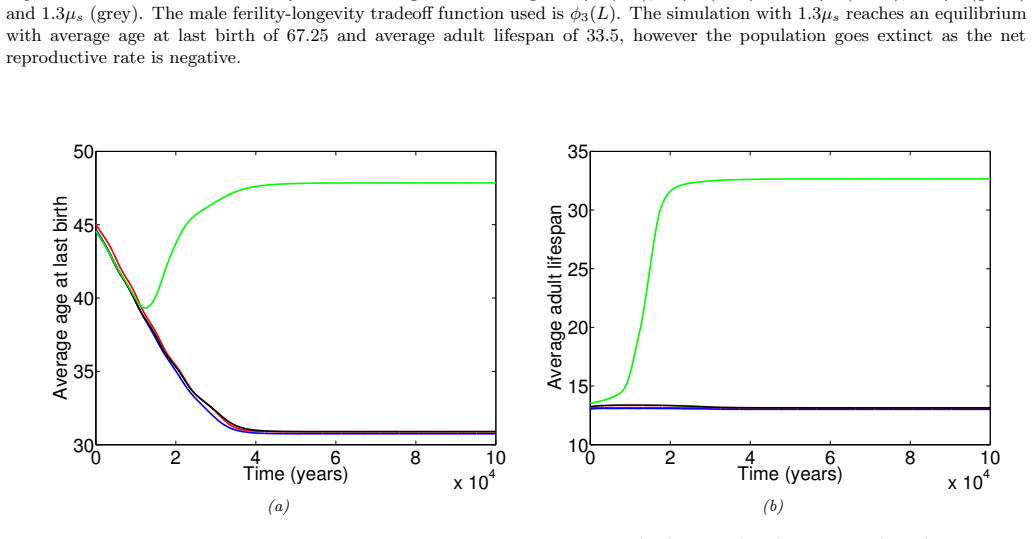
In the extended two-sex PDE model, grandmothering restricted to daughters' fertility produces only two locally stable equilibria for longevity and reproductive span: one matching great-ape life histories and one matching hunter-gatherer life histories. The grandmothering effect permits the transition between these equilibria without any extension of the age at last birth. Sensitivity analysis shows that the strength of male competition, set by mating sex-ratio skew, determines both whether the transition to higher longevity is reachable and the equilibrium value of average adult lifespan.
What carries the argument
The two-sex partial differential equation that couples evolving adult longevity, age at last birth, grandmothering support to daughters, and male competition driven by mating sex-ratio skew.
If this is right
- Grandmothering alone is sufficient to produce the observed increase in human longevity from ape-like baselines.
- The equilibrium adult lifespan depends on the intensity of male competition.
- The equilibrium age at last birth is shifted by grandmothering while adult lifespan is not.
- No change in the physiological end of fertility is required for the longevity increase.
Where Pith is reading between the lines
- Social rules that channel grandmother help exclusively to daughters may be necessary for the longevity transition to remain stable.
- Changes in mating systems that alter sex-ratio skew could block or enable similar longevity shifts in other species.
- The model predicts that populations with strong grandmothering should show earlier age at last birth than expected from longevity alone.
Load-bearing premise
Post-fertile females provide reproductive support only to their daughters and male competition is driven solely by the mating sex-ratio skew.
What would settle it
A simulation or empirical comparison in which grandmothering support is extended to sons or other relatives and the two equilibria merge or the transition to higher longevity disappears.
Figures
read the original abstract
We use a two-sex partial differential equation (PDE) model based on the Grandmother Hypothesis. We build on an earlier model by Kim et al. [27] by allowing for evolution in both longevity and age at last birth, and also assuming that post-fertile females support only their daughters' fertility. Similarly to Kim et al. [27], we find that only two locally stable equilibria exist: one corresponding to great ape-like longevities and the other corresponding to hunter-gatherer longevities. Our results show that grandmothering enables the transition between these two equilibria, without extending the end of fertility. Moreover, sensitivity analyses of the model show that male competition, arising from a skew in the mating sex ratio towards males, plays a significant role in determining whether the transition from great ape-like longevities to higher longevities is possible and the equilibrium value of the average adult lifespan. Whereas grandmothering effects have a significant impact on the equilibrium value of the average age at last birth and enable the transition to higher longevities, they have an insignificant impact on the equilibrium value of the average adult lifespan.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript presents a two-sex PDE model extending the Grandmother Hypothesis from Kim et al. (2019), allowing evolution in both longevity and age at last birth while restricting post-fertile female support to daughters' fertility only. It reports that only two locally stable equilibria exist—one with great ape-like longevities and one with hunter-gatherer longevities—with grandmothering enabling the transition between them without extending the end of fertility. Sensitivity analyses indicate that male competition (via mating sex ratio skew) significantly affects transition feasibility and equilibrium adult lifespan, while grandmothering strongly affects equilibrium age at last birth but not adult lifespan.
Significance. If the equilibria and transition results hold under the stated assumptions, the work provides a mechanistic account of how grandmothering can shift populations from ape-like to human-like longevity without fertility extension, while quantifying the modulating role of male competition. The use of a two-sex PDE framework and sensitivity analyses on the mating skew parameter are strengths that allow exploration of sexual conflict effects.
major comments (2)
- [Abstract and model description] The central claim that only two locally stable equilibria exist (and that grandmothering enables the transition) is load-bearing but presented without the explicit PDE system, functional forms for fertility/mortality, or the numerical method used to locate and classify the equilibria. This prevents independent verification that the reported equilibria are reached from realistic initial conditions.
- [Sensitivity analyses] The sensitivity results on male competition and grandmothering strength are reported without specific parameter values, ranges, or error analysis (e.g., how the mating sex ratio skew parameter and grandmothering support strength to daughters are varied). This makes it impossible to assess the robustness of the claim that male competition determines transition possibility.
Simulated Author's Rebuttal
We thank the referee for their constructive and detailed comments. We address each major comment below and have made revisions to improve clarity and verifiability of the results.
read point-by-point responses
-
Referee: [Abstract and model description] The central claim that only two locally stable equilibria exist (and that grandmothering enables the transition) is load-bearing but presented without the explicit PDE system, functional forms for fertility/mortality, or the numerical method used to locate and classify the equilibria. This prevents independent verification that the reported equilibria are reached from realistic initial conditions.
Authors: The explicit two-sex PDE system, functional forms for fertility and mortality (including the grandmothering term restricted to daughters), and the numerical methods (finite-difference discretization, continuation to locate equilibria, and linear stability analysis) are fully specified in the Methods section, along with the initial conditions corresponding to ape-like demography. To make these details more immediately accessible and address the verification concern, we will add a concise presentation of the core PDE equations and numerical approach to the main text in the revised manuscript. revision: yes
-
Referee: [Sensitivity analyses] The sensitivity results on male competition and grandmothering strength are reported without specific parameter values, ranges, or error analysis (e.g., how the mating sex ratio skew parameter and grandmothering support strength to daughters are varied). This makes it impossible to assess the robustness of the claim that male competition determines transition possibility.
Authors: The parameter ranges and variation protocol for the mating-sex-ratio skew (varied systematically from 1 to 4) and grandmothering support strength (varied from 0 to 0.5) are documented in the supplementary materials and associated table. In the revision we will add an explicit subsection in Results that states these ranges, the discrete steps used, the criterion for declaring a transition feasible, and any robustness checks performed across initial conditions. revision: yes
Circularity Check
No significant circularity in derivation chain
full rationale
The paper extends an existing two-sex PDE model from the cited Kim et al. reference by adding evolution in both longevity and age at last birth plus a restriction on grandmothering support. The reported finding of two locally stable equilibria is obtained by direct analysis of the extended system; the equilibria are not obtained by fitting parameters to data and relabeling the fit as a prediction, nor does any load-bearing step reduce to a self-citation that substitutes for an independent derivation. The functional forms and structural choices are carried forward explicitly as modeling decisions, but the extension itself supplies independent content within the stated framework. No self-definitional, fitted-input, or ansatz-smuggling patterns are present in the abstract or described claims.
Axiom & Free-Parameter Ledger
free parameters (2)
- mating sex ratio skew parameter
- grandmothering support strength to daughters
axioms (2)
- domain assumption Post-fertile females support only their daughters' fertility
- standard math Two-sex PDE system with continuous age and time
Reference graph
Works this paper leans on
-
[1]
Alberts, S. C., Altmann, J., Brockman, D. K., Cords, M., Fedigan, L. M., Pusey, A., Stoinski, T. S., Strier, K. B., Morris, W. F., and Bronikowski, A. M. (2013). Reproductive aging patterns in primates reveal that humans are distinct. Proc. Natl. Acad. Sci. U.S.A. , 110(33):13440–13445
work page 2013
-
[2]
Arnqvist, G. and Rowe, L. (2005). Sexual Conflict . Monographs in behavior and ecology. Princeton University Press
work page 2005
-
[3]
G., Hawkes, K., and O’Connell, J
Blurton Jones, N. G., Hawkes, K., and O’Connell, J. F. (2002). Antiquity of post-reproductive life: are there modern impacts on hunter-gatherer post-reproductive life spans? American Journal of Human Biology , 14(2):184– 205
work page 2002
-
[4]
Boesch, C., Kohou, G., Nene, H., and Vigilant, L. (2006). Male competition and paternity in wild chimpanzees of the Ta¨ ı forest.American Journal of Physical Anthropology, 130(1):103–115
work page 2006
-
[5]
Charnov, E. L. (1993). Life History Invariants: Some Explorations of Symmetry in Evolutionary Ecology . Life History Invariants: Some Explorations of Symmetry in Evolutionary Ecology. Oxford University Press
work page 1993
-
[6]
Coale, A. and Demeny, P. (1968). Regional Model Life Tables and Stable Population . Princeton University Press, New Jersey
work page 1968
-
[7]
Coxworth, J. E., Kim, P. S., McQueen, J. S., and Hawkes, K. (2015). Grandmothering life histories and human pair bonding. Proceedings of the National Academy of Sciences , 112(38):11806–11811
work page 2015
-
[8]
Croft, D. P., Brent, L. J., Franks, D. W., and Cant, M. A. (2015). The evolution of prolonged life after reproduction. Trends in Ecology and Evolution , 30(7):407–416
work page 2015
-
[9]
Field, J. M., Hawkes, K., and Kim, P. S. (2015). A continuous model of grandmothering: thresholds, evolutionary stability, and who gets help. In review
work page 2015
-
[10]
Gardner, A. and West, S. A. (2006). Demography, altruism, and the benefits of budding. Journal of Evolutionary Biology, 19(5):1707–1716
work page 2006
-
[11]
Gurven, M. and Kaplan, H. (2007). Longevity among hunter-gatherers: A cross-cultural examination. Population and Development Review , 33(2):321–365
work page 2007
-
[12]
Hamilton, W. D. (1964a). The genetical evolution of social behaviour. I. Journal of Theoretical Biology, 7(1):1–16
-
[13]
Hamilton, W. D. (1964b). The genetical evolution of social behaviour. II. Journal of Theoretical Biology, 7(1):17– 52
-
[14]
Hawkes, K. (2003). Grandmothers and the evolution of human longevity. American Journal of Human Biology , 15(3):380–400
work page 2003
-
[15]
Hawkes, K. (2004). Human longevity: the grandmother effect. Nature, 428(6979):128–129
work page 2004
-
[16]
S., Kennedy, B., Bohlender, R., and Hawks, J
Hawkes, K., Kim, P. S., Kennedy, B., Bohlender, R., and Hawks, J. (2011). A reappraisal of grandmothering and natural selection. Proceedings of the Royal Society of London B: Biological Sciences , 278(1714):1936–1938
work page 2011
-
[17]
Hawkes, K., O’Connell, J. F., and Blurton Jones, N. G. (1995). Hadza children’s foraging: Juvenile dependency, social arrangements, and mobility among hunter-gatherers. Current Anthropology, 36(4):688–700
work page 1995
-
[18]
Hawkes, K., O’Connell, J. F., and Blurton Jones, N. G. (1997). Hadza women’s time allocation, offspring provisioning, and the evolution of long postmenopausal life spans. Current Anthropology, 38(4):551–577
work page 1997
-
[19]
Hawkes, K., O’Connell, J. F., Blurton Jones, N. G., Alvarez, H., and Charnov, E. L. (1998). Grandmothering, menopause, and the evolution of human life histories. Proceedings of the National Academy of Sciences, 95(3):1336– 1339. 20
work page 1998
-
[20]
Hawkes, K., Smith, K. R., and Robson, S. L. (2009). Mortality and fertility rates in humans and chimpanzees: How within-species variation complicates cross-species comparisons. American Journal of Human Biology, 21(4):578–586
work page 2009
-
[21]
Hill, K., Boesch, C., Goodall, J., Pusey, A., Williams, J., and Wrangham, R. (2001). Mortality rates among wild chimpanzees. Journal of Human Evolution , 40(5):437–450
work page 2001
-
[22]
Hill, K. and Hurtado, A. (1996). Ache life history: The ecology and demography of a foraging people . Aldine de Gruyter, New York
work page 1996
-
[23]
Howell, N. (1979). Demography of the Dobe !Kung . Academic Press, New York, 1 edition
work page 1979
-
[24]
Kachel, A. F., Premo, L. S., and Hublin, J. (2010). Grandmothering and natural selection. Proceedings of the Royal Society of London B: Biological Sciences
work page 2010
-
[25]
Kachel, A. F., Premo, L. S., and Hublin, J. (2011). Grandmothering and natural selection revisited. Proceedings of the Royal Society of London B: Biological Sciences , 278(1714):1939–1941
work page 2011
-
[26]
Kim, P. S., Coxworth, J. E., and Hawkes, K. (2012). Increased longevity evolves from grandmothering.Proceedings of the Royal Society of London B: Biological Sciences
work page 2012
-
[27]
Kim, P. S., McQueen, J. S., Coxworth, J. E., and Hawkes, K. (2014). Grandmothering drives the evolution of longevity in a probabilistic model. Journal of Theoretical Biology, 353:84–94
work page 2014
-
[28]
Kokko, H. and Jennions, M. D. (2014). The relationship between sexual selection and sexual conflict. Cold Spring Harbor Perspectives in Biology, 6(9):a017517
work page 2014
-
[29]
Lahdenper¨ o, M., Gillespie, D. O. S., Lummaa, V., and Russell, A. F. (2012). Severe intergenerational reproductive conflict and the evolution of menopause. Ecology Letters, 15(11):1283–1290
work page 2012
-
[30]
Lee, R. (2008). Sociality, selection, and survival: Simulated evolution of mortality with intergenerational transfers and food sharing. Proceedings of the National Academy of Sciences , 105(20):7124–7128
work page 2008
-
[31]
Levitis, D. A., Burger, O., and Lackey, L. B. (2013). The human post-fertile lifespan in comparative evolutionary context. Evolutionary Anthropology, 22(2):66–79
work page 2013
-
[32]
Levitis, D. A. and Lackey, L. B. (2011). A measure for describing and comparing post-reproductive lifespan as a population trait. Methods in Ecology and Evolution , 2(5):446–453
work page 2011
-
[33]
Mitteldorf, J. and Wilson, D. S. (2000). Population viscosity and the evolution of altruism. Journal of Theoretical Biology, 204(4):481–496
work page 2000
-
[34]
F., Hawkes, K., and Blurton Jones, N
O’Connell, J. F., Hawkes, K., and Blurton Jones, N. G. (1999). Grandmothering and the evolution of homo erectus. Journal of Human Evolution , 36(5):461 – 485
work page 1999
-
[35]
Pavard, S. and Branger, F. (2012). Effect of maternal and grandmaternal care on population dynamics and human life-history evolution: a matrix projection model. Theor Popul Biol , 82(4):364–376
work page 2012
-
[36]
Peccei, J. S. (2001). Menopause: Adaptation or epiphenomenon? Evolutionary Anthropology: Issues, News, and Reviews, 10(2):43–57
work page 2001
-
[37]
Robbins, M. M. (1995). A demographic analysis of male life history and social structure of mountain gorillas. Behaviour, 132(1/2):21–47
work page 1995
-
[38]
Sear, R. and Mace, R. (2008). Who keeps children alive? a review of the effects of kin on child survival. Evolution and Human Behavior , 29(1):1–18
work page 2008
-
[39]
Sear, R., Mace, R., and McGregor, I. A. (2000). Maternal grandmothers improve nutritional status and survival of children in rural gambia. Proceedings of the Royal Society of London B: Biological Sciences, 267(1453):1641–1647
work page 2000
-
[40]
Sugiyama, Y. (1994). Age-specific birth rate and lifetime reproductive success of chimpanzees at bossou, guinea. American Journal of Primatology , 32(4):311–318. 21
work page 1994
-
[41]
Taylor, P. (1992a). Altruism in viscous populations – an inclusive fitness model. Evolutionary Ecology, 6(4):352– 356
-
[42]
Taylor, P. (1992b). Inclusive Fitness in a Homogeneous Environment. Proceedings: Biological Sciences , 249(1326):299–302
-
[43]
te Velde, E. R. and Pearson, P. L. (2002). The variability of female reproductive ageing. Human Reproduction Update, 8(2):141–154
work page 2002
-
[44]
M., Kaplan, H., Kramer, K., Oliver, W
Walker, R., Gurven, M., Hill, K., Migliano, A., Chagnon, N., De Souza, R., Djurovic, G., Hames, R., Hurtado, A. M., Kaplan, H., Kramer, K., Oliver, W. J., Valeggia, C., and Yamauchi, T. (2006). Growth rates and life histories in twenty-two small-scale societies. American Journal of Human Biology , 18(3):295–311
work page 2006
-
[45]
West, S. A., Pen, I., and Griffin, A. S. (2002). Cooperation and competition between relatives. Science, 296(5565):72–75
work page 2002
-
[46]
Williams, G. C. (1957). Pleiotropy, natural selection, and the evolution of senescence. Evolution, 11(4):398–411
work page 1957
-
[47]
Wilson, D. S., Pollock, G. B., and Dugatkin, L. A. (1992). Can altruism evolve in purely viscous populations? Evolutionary Ecology, 6(4):331–341. 22
work page 1992
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.