Matters of Life and Death in Computational Cell Biology
Pith reviewed 2026-05-21 20:06 UTC · model grok-4.3
The pith
Computational cell biology requires centering the life-death boundary to create a theory of cellular viability.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
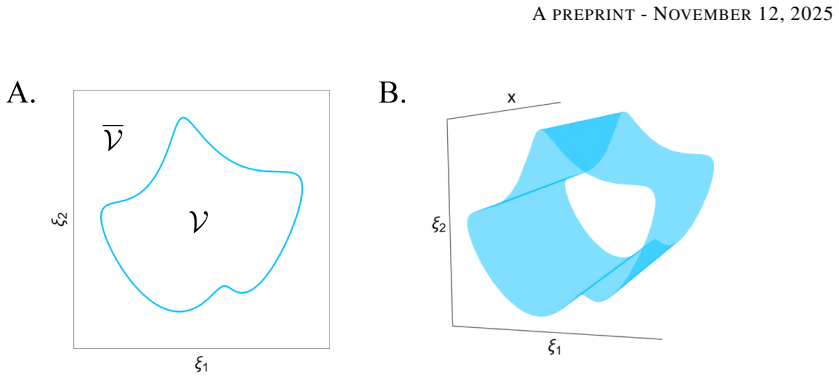
Treating the life-death boundary as a central concept in computational cell models reveals specific geometric structures that separate regions of qualitatively similar survival outcomes, thereby supplying new global organizing principles for cell fate.
What carries the argument
Geometric structures in model space that partition regions of similar survival outcomes, generated once the life-death boundary is placed at the center of analysis.
If this is right
- Cell fate decisions gain a geometric description rather than depending on ad hoc constraint checks.
- Idealized models of emergent individuals become the starting point for mapping intrinsic viability limits.
- Biophysical constraints acquire a unified framework instead of remaining case-by-case additions.
Where Pith is reading between the lines
- The same geometric approach could guide which quantities experimental biologists measure near viability thresholds.
- Links may appear to broader questions of self-maintenance in other complex dynamical systems.
- Extending the method to higher-resolution cell models would test whether the geometric patterns generalize.
Load-bearing premise
Biophysical constraints are implemented without systematicity in cell models, and centering the life-death boundary will produce a workable theory of cellular viability.
What would settle it
Demonstrating that geometric structures fail to separate regions of similar survival outcomes across multiple cell models would undermine the proposed organizing principles.
Figures





read the original abstract
Nearly all cell models explicitly or implicitly deal with the biophysical constraints that must be respected for life to persist. Despite this, there is almost no systematicity in how these constraints are implemented, and we lack a principled understanding of how cellular dynamics interact with them and how they originate in actual biology. Computational cell biology will only overcome these concerns once it treats the life-death boundary as a central concept, creating a theory of cellular viability. We lay the foundation for such a development by demonstrating how specific geometric structures can separate regions of qualitatively similar survival outcomes in our models, offering new global organizing principles for cell fate. We also argue that idealized models of emergent individuals offer a tractable way to begin understanding life's intrinsically generated limits.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript argues that computational cell biology lacks systematicity in implementing biophysical constraints for cell survival despite their implicit presence in models. It proposes treating the life-death boundary as a central concept to develop a theory of cellular viability. The authors claim to lay the foundation for this by demonstrating how specific geometric structures separate regions of qualitatively similar survival outcomes in their models, thereby offering new global organizing principles for cell fate. They further suggest that idealized models of emergent individuals provide a tractable route to understanding life's intrinsically generated limits.
Significance. If the geometric separation of survival regions can be rigorously shown in concrete models and generalized, the work could supply useful organizing principles that help systematize viability constraints across computational cell biology. This framing might encourage more unified approaches to modeling cell fate under biophysical limits, though its impact hinges on providing verifiable technical content rather than remaining at the conceptual level.
major comments (2)
- [Abstract] Abstract: The claim that 'specific geometric structures can separate regions of qualitatively similar survival outcomes in our models' is asserted without model equations, state-space definitions, parameter regimes, simulation protocols, or figures illustrating the claimed partitions. This absence makes the central demonstration load-bearing for the paper's foundation-laying assertion unverifiable from the provided content.
- [Main text] Main argument: The geometric demonstration relies exclusively on the authors' own models without external benchmarks, independent validation, or comparison to existing cell-fate frameworks, which risks circularity in defining the 'new global organizing principles' in terms of the very models under analysis.
minor comments (1)
- [Abstract] The repeated use of 'our models' without prior specification of which models or their key assumptions reduces clarity for readers unfamiliar with the authors' prior work.
Simulated Author's Rebuttal
We thank the referee for their constructive report and for recognizing the potential value of centering the life-death boundary in computational cell biology. We address each major comment below, indicating where revisions will be made to strengthen the manuscript.
read point-by-point responses
-
Referee: [Abstract] Abstract: The claim that 'specific geometric structures can separate regions of qualitatively similar survival outcomes in our models' is asserted without model equations, state-space definitions, parameter regimes, simulation protocols, or figures illustrating the claimed partitions. This absence makes the central demonstration load-bearing for the paper's foundation-laying assertion unverifiable from the provided content.
Authors: We acknowledge that the abstract presents the core claim at a conceptual level without the supporting technical details. The full manuscript contains descriptions of the models and geometric analyses, but to improve verifiability we will add a concise methods subsection summarizing the key equations, state-space definitions, and parameter regimes, together with a new figure that explicitly illustrates the partitioned survival regions for at least one representative model. revision: yes
-
Referee: [Main text] Main argument: The geometric demonstration relies exclusively on the authors' own models without external benchmarks, independent validation, or comparison to existing cell-fate frameworks, which risks circularity in defining the 'new global organizing principles' in terms of the very models under analysis.
Authors: The models function as concrete, minimal illustrations of how geometric structures arise from biophysical constraints rather than as the sole source of the organizing principles. The principles themselves are framed as general features of viability boundaries that can be examined in any model respecting conservation laws and energy dissipation. To address the concern about circularity we will expand the discussion section to compare the geometric approach with established cell-fate frameworks (e.g., Boolean network models of apoptosis and continuous differential-equation descriptions of metabolic thresholds), thereby situating the contribution relative to the existing literature. revision: partial
Circularity Check
No significant circularity; conceptual proposal remains self-contained
full rationale
The paper is framed as a position piece that identifies a lack of systematicity in biophysical constraints across cell models and proposes treating the life-death boundary centrally, with a demonstration of geometric structures claimed in the authors' models. No load-bearing derivation chain reduces a result to its own inputs by construction: there are no equations shown that define a quantity in terms of itself, no fitted parameters renamed as predictions, and no uniqueness theorems or ansatzes imported solely via self-citation that forbid alternatives. The central claim is presented as an organizing principle derived from the models rather than tautologically equivalent to them, and the argument does not rely on a self-referential loop for its validity. This is the normal finding for a conceptual manuscript without explicit self-referential reductions in its technical steps.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption Nearly all cell models explicitly or implicitly deal with biophysical constraints that must be respected for life to persist.
- domain assumption There is almost no systematicity in how these constraints are implemented and a lack of principled understanding of their interaction with cellular dynamics.
Reference graph
Works this paper leans on
-
[1]
Opportunities and challenges for digital twins in biomedical research: Proceedings of a workshop-in brief, 2023. Pages: 26922
work page 2023
- [2]
- [3]
-
[4]
E. Agmon and R. K. Spangler. A multi-scale approach to modeling e. coli chemotaxis. Entropy, 22(10):1101, 2020
work page 2020
-
[5]
E. Agmon and B. R. Stockwell. Lipid homeostasis and regulated cell death. Current Opinion in Chemical Biology, 39:83–89, 2017
work page 2017
-
[6]
K. Aihara and H. Suzuki. Theory of hybrid dynamical systems and its applications to biological and medical systems. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences, 368(1930):4893–4914, 2010
work page 1930
-
[7]
A. R. A. Anderson, K. A. Rejniak, P. Gerlee, and V . Quaranta. Microenvironment driven invasion: a multiscale multimodel investigation. Journal of Mathematical Biology, 58(4):579–624, 2009
work page 2009
-
[8]
W. R. Ashby. Design for a Brain. Springer Netherlands, 1960
work page 1960
- [9]
-
[10]
X. E. Barandiaran and M. D. Egbert. Norm-establishing and norm-following in autonomous agency. Artificial Life, 20(1):5–28, 2014
work page 2014
- [11]
-
[12]
S. Bartlett, A. W. Eckford, M. Egbert, M. Lingam, A. Kolchinsky, A. Frank, and G. Ghoshal. Physics of life: Exploring information as a distinctive feature of living systems. PRX Life, 3(3):037003, 2025
work page 2025
-
[13]
R. D. Beer. A dynamical systems perspective on agent-environment interaction. Artificial Intelligence, 72(1):173– 215, 1995
work page 1995
-
[14]
R. D. Beer. Characterizing autopoiesis in the game of life. Artificial Life, 21(1):1–19, 2015
work page 2015
-
[15]
R. D. Beer. An integrated perspective on the constitutive and interactive dimensions of autonomy. In The 2020 Conference on Artificial Life, pages 202–209. MIT Press, 2020
work page 2020
-
[16]
R. D. Beer, A. Barwich, and G. J. Severino. Milking a spherical cow: Toy models in neuroscience. European Journal of Neuroscience, 60(10):6359–6374, 2024
work page 2024
-
[17]
R. D. Beer and E. A. Di Paolo. The theoretical foundations of enaction: Precariousness. Biosystems, 223:104823, 2023
work page 2023
-
[18]
R. D. Beer, C. McShaffrey, and T. M. Gaul. Deriving the intrinsic viability constraint of an emergent individual from first principles. In The 2024 Conference on Artificial Life. MIT Press, 2024
work page 2024
-
[19]
R. D. Beer and P. L. Williams. Animals and animats: Why not both iguanas? Adaptive Behavior, 17(4):296–302, 2009
work page 2009
- [20]
-
[21]
T. Bergsbaken, S. L. Fink, and B. T. Cookson. Pyroptosis: host cell death and inflammation. Nature Reviews Microbiology, 7(2):99–109, 2009
work page 2009
-
[22]
F. J. Bock and S. W. G. Tait. Mitochondria as multifaceted regulators of cell death.Nature Reviews Molecular Cell Biology, 21(2):85–100, 2020
work page 2020
-
[23]
B. W.-C. Chan. Lenia - biology of artificial life. 2018. Publisher: arXiv Version Number: 3
work page 2018
-
[24]
B. W.-C. Chan. Lenia and expanded universe. In The 2020 Conference on Artificial Life, pages 221–229. MIT Press, 2020
work page 2020
-
[25]
H. Cho, T. Uehara, and T. Bernhardt. Beta-lactam antibiotics induce a lethal malfunctioning of the bacterial cell wall synthesis machinery. Cell, 159(6):1300–1311, 2014
work page 2014
-
[26]
A. Ciccia and S. J. Elledge. The DNA damage response: Making it safe to play with knives. Molecular Cell, 40(2):179–204, 2010
work page 2010
- [27]
-
[28]
J. D. Davis, C. M. Kumbale, Q. Zhang, and E. O. V oit. Dynamical systems approaches to personalized medicine. Current Opinion in Biotechnology, 58:168–174, 2019
work page 2019
- [29]
-
[30]
P. Dittrich, J. Ziegler, and W. Banzhaf. Artificial chemistries—a review. Artificial Life, 7(3):225–275, 2001
work page 2001
-
[31]
M. D. Egbert and J. Pérez-Mercader. Methods for measuring viability and evaluating viability indicators.Artificial Life, 24(2):106–118, 2018
work page 2018
-
[32]
T. Eissing, H. Conzelmann, E. D. Gilles, F. Allgöwer, E. Bullinger, and P. Scheurich. Bistability analyses of a caspase activation model for receptor-induced apoptosis. Journal of Biological Chemistry, 279(35):36892–36897, 2004
work page 2004
-
[33]
S. Elmore. Apoptosis: A review of programmed cell death. Toxicologic Pathology, 35(4):495–516, 2007
work page 2007
-
[34]
M. Elowitz and W. A. Lim. Build life to understand it. Nature, 468(7326):889–890, 2010
work page 2010
-
[35]
K. M. Evans. Larger than life: Digital creatures in a family of two-dimensional cellular automata. Discrete Mathematics & Theoretical Computer Science, DMTCS Proceedings vol. AA,...:2288, 2001
work page 2001
-
[36]
J. Ferrell. Bistability, bifurcations, and waddington’s epigenetic landscape. Current Biology, 22(11):R458–R466, 2012
work page 2012
-
[37]
R. A. Gatenby, K. Smallbone, P. K. Maini, F. Rose, J. Averill, R. B. Nagle, L. Worrall, and R. J. Gillies. Cellular adaptations to hypoxia and acidosis during somatic evolution of breast cancer. British Journal of Cancer , 97(5):646–653, 2007
work page 2007
-
[38]
T. M. Gaul. Autopoiesis in RealLife euclidean automata. In The 2024 Conference on Artificial Life. MIT Press, 2024
work page 2024
-
[39]
A. Ghaffarizadeh, R. Heiland, S. H. Friedman, S. M. Mumenthaler, and P. Macklin. PhysiCell: An open source physics-based cell simulator for 3-d multicellular systems. PLOS Computational Biology, 14(2):e1005991, 2018
work page 2018
-
[40]
P. Ghose and S. Shaham. Cell death in animal development. Development, 147(14):dev191882, 2020
work page 2020
- [41]
-
[42]
Y . Hao, S. Hao, E. Andersen-Nissen, W. M. Mauck, S. Zheng, A. Butler, M. J. Lee, A. J. Wilk, C. Darby, M. Zager, P. Hoffman, M. Stoeckius, E. Papalexi, E. P. Mimitou, J. Jain, A. Srivastava, T. Stuart, L. M. Fleming, B. Yeung, A. J. Rogers, J. M. McElrath, C. A. Blish, R. Gottardo, P. Smibert, and R. Satija. Integrated analysis of multimodal single-cell ...
work page 2021
-
[43]
Y . Himeoka, S. A. Horiguchi, and T. J. Kobayashi. Theoretical basis for cell deaths.Physical Review Research, 6(4):043217, 2024
work page 2024
-
[44]
G. T. Johnson, E. Agmon, M. Akamatsu, E. Lundberg, B. Lyons, W. Ouyang, O. A. Quintero-Carmona, M. Riel- Mehan, S. Rafelski, and R. Horwitz. Building the next generation of virtual cells to understand cellular biology. Biophysical Journal, 122(18):3560–3569, 2023
work page 2023
-
[45]
O. Karin and U. Alon. Biphasic response as a mechanism against mutant takeover in tissue homeostasis circuits. Molecular Systems Biology, 13(6):933, 2017
work page 2017
-
[46]
J. Karr, J. Sanghvi, D. Macklin, M. Gutschow, J. Jacobs, B. Bolival, N. Assad-Garcia, J. Glass, and M. Covert. A whole-cell computational model predicts phenotype from genotype. Cell, 150(2):389–401, 2012
work page 2012
-
[47]
K. Kochanowski, H. Okano, V . Patsalo, J. Williamson, U. Sauer, and T. Hwa. Global coordination of metabolic pathways in Escherichia coli by active and passive regulation. Molecular Systems Biology, 17(4):e10064, 2021
work page 2021
-
[48]
A. Kolchinsky and D. H. Wolpert. Semantic information, autonomous agency and non-equilibrium statistical physics. Interface Focus, 8(6):20180041, 2018
work page 2018
- [49]
-
[50]
J. T. Lennon and S. E. Jones. Microbial seed banks: the ecological and evolutionary implications of dormancy. Nature Reviews Microbiology, 9(2):119–130, 2011
work page 2011
-
[51]
X. Li, P. Zhang, Z. Yin, F. Xu, Z.-H. Yang, J. Jin, J. Qu, Z. Liu, H. Qi, C. Yao, and J. Shuai. Caspase-1 and gasdermin d afford the optimal targets with distinct switching strategies in NLRP1b inflammasome-induced cell death. Research, 2022:2022/9838341, 2022
work page 2022
-
[52]
J. S. Lowengrub, H. B. Frieboes, F. Jin, Y .-L. Chuang, X. Li, P. Macklin, S. M. Wise, and V . Cristini. Nonlinear modelling of cancer: Bridging the gap between cells and tumours. Nonlinearity, 23(1):R1–R91, 2010
work page 2010
-
[53]
P. Macklin, S. Mumenthaler, and J. Lowengrub. Modeling multiscale necrotic and calcified tissue biomechanics in cancer patients: Application to ductal carcinoma in situ (DCIS). In A. Gefen, editor, Multiscale Computer Modeling in Biomechanics and Biomedical Engineering, volume 14, pages 349–380. Springer Berlin Heidelberg,
-
[54]
12 A PREPRINT - NOVEMBER 12, 2025
Series Title: Studies in Mechanobiology, Tissue Engineering and Biomaterials. 12 A PREPRINT - NOVEMBER 12, 2025
work page 2025
- [55]
-
[56]
G. Majno and I. Joris. Apoptosis, oncosis, and necrosis. an overview of cell death. The American Journal of Pathology, 146(1):3–15, 1995
work page 1995
-
[57]
H. R. Maturana and F. J. Varela. Autopoiesis and cognition: the realization of the living. Boston studies in the philosophy of science. D. Reidel Publishing Company, 1980
work page 1980
-
[58]
C. McShaffrey and R. D. Beer. Decomposing viability space. In The 2023 Conference on Artificial Life. MIT Press, 2023
work page 2023
-
[59]
C. McShaffrey and R. D. Beer. Dissecting viability in multi-agent systems. In The 2024 Conference on Artificial Life. MIT Press, 2024
work page 2024
-
[60]
C. McShaffrey and R. D. Beer. Shaking up viability space: Stochasticity and survival in the transient. In The 2025 Conference on Artificial Life. MIT Press, 2025
work page 2025
-
[61]
C. McShaffrey and R. D. Beer. Viability space: A geometric decomposition of survival outcomes in single- and multi-agent systems. In Preparation
-
[62]
J. D. Orth, I. Thiele, and B. Ø. Palsson. What is flux balance analysis? Nature Biotechnology, 28(3):245–248, 2010
work page 2010
-
[63]
M. Pasparakis and P. Vandenabeele. Necroptosis and its role in inflammation. Nature, 517(7534):311–320, 2015
work page 2015
- [64]
-
[65]
M. Rehm, H. J. Huber, H. Dussmann, and J. H. M. Prehn. Systems analysis of effector caspase activation and its control by x-linked inhibitor of apoptosis protein. The EMBO Journal, 25(18):4338–4349, 2006
work page 2006
-
[66]
D. H. Rothman and S. Zaleski. Lattice-gas models of phase separation: interfaces, phase transitions, and multiphase flow. Reviews of Modern Physics, 66(4):1417–1479, 1994
work page 1994
-
[67]
M. Ruscone, A. Checcoli, R. Heiland, E. Barillot, P. Macklin, L. Calzone, and V . Noël. Building multiscale models with PhysiBoSS, an agent-based modeling tool. Briefings in Bioinformatics, 25(6):bbae509, 2024
work page 2024
-
[68]
A. Régimbeau, M. Budinich, A. Larhlimi, J. J. Pierella Karlusich, O. Aumont, L. Memery, C. Bowler, and D. Eveillard. Contribution of genome-scale metabolic modelling to niche theory. Ecology Letters, 25(6):1352– 1364, 2022
work page 2022
- [69]
-
[70]
M. R. Shaebani, A. Wysocki, R. G. Winkler, G. Gompper, and H. Rieger. Computational models for active matter. Nature Reviews Physics, 2(4):181–199, 2020
work page 2020
-
[71]
C. J. Skalnik, E. Agmon, R. K. Spangler, L. Talman, J. H. Morrison, S. M. Peirce, and M. W. Covert. Whole-colony modeling of escherichia coli, 2021
work page 2021
-
[72]
K. Smallbone, R. A. Gatenby, R. J. Gillies, P. K. Maini, and D. J. Gavaghan. Metabolic changes during carcinogenesis: Potential impact on invasiveness. Journal of Theoretical Biology, 244(4):703–713, 2007
work page 2007
-
[73]
R. Solé, C. P. Kempes, B. Corominas-Murtra, M. De Domenico, A. Kolchinsky, M. Lachmann, E. Libby, S. Saavedra, E. Smith, and D. Wolpert. Fundamental constraints to the logic of living systems. Interface Focus, 14(5):20240010, 2024
work page 2024
-
[74]
S. Spencer and P. Sorger. Measuring and modeling apoptosis in single cells. Cell, 144(6):926–939, 2011
work page 2011
-
[75]
J. Starruß, W. De Back, L. Brusch, and A. Deutsch. Morpheus: a user-friendly modeling environment for multiscale and multicellular systems biology. Bioinformatics, 30(9):1331–1332, 2014
work page 2014
- [76]
-
[77]
M. H. Swat, G. L. Thomas, J. M. Belmonte, A. Shirinifard, D. Hmeljak, and J. A. Glazier. Multi-scale modeling of tissues using CompuCell3d. In Methods in Cell Biology, volume 110, pages 325–366. Elsevier, 2012
work page 2012
-
[78]
B. E. Trump, I. K. Berezesky, S. H. Chang, and P. C. Phelps. The pathways of cell death: Oncosis, apoptosis, and necrosis. Toxicologic Pathology, 25(1):82–88, 1997
work page 1997
-
[79]
F. J. Varela. Principles of biological autonomy. The MIT Press, new annotated edition edition, 2025
work page 2025
-
[80]
E. O. V oit. A systems-theoretical framework for health and disease: Inflammation and preconditioning from an abstract modeling point of view. Mathematical Biosciences, 217(1):11–18, 2009. 13 A PREPRINT - NOVEMBER 12, 2025
work page 2009
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.