Two-population model of type Ia supernovae and their associations with host galaxies in ZTF DR2
Pith reviewed 2026-06-26 23:48 UTC · model grok-4.3
The pith
Two-population model of type Ia supernovae makes host-galaxy step corrections redundant.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
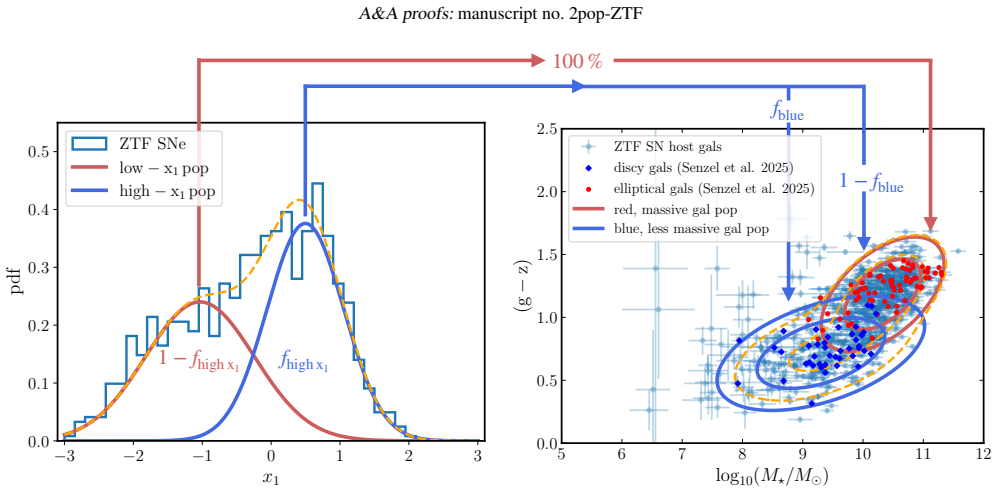
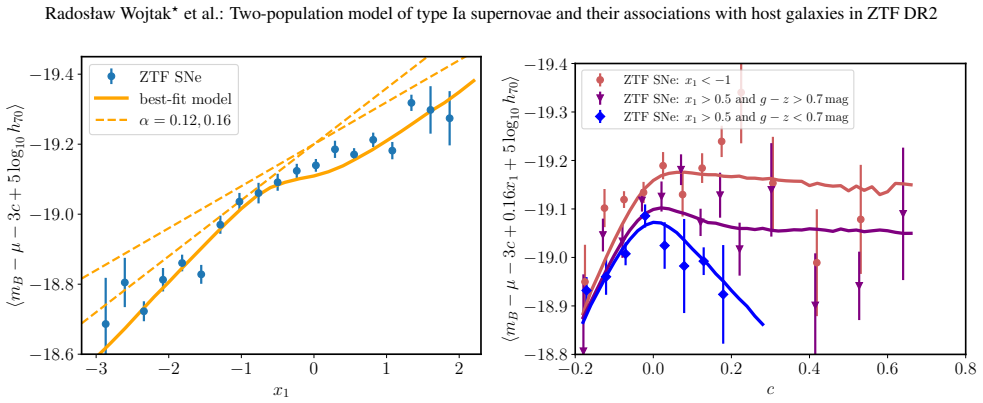
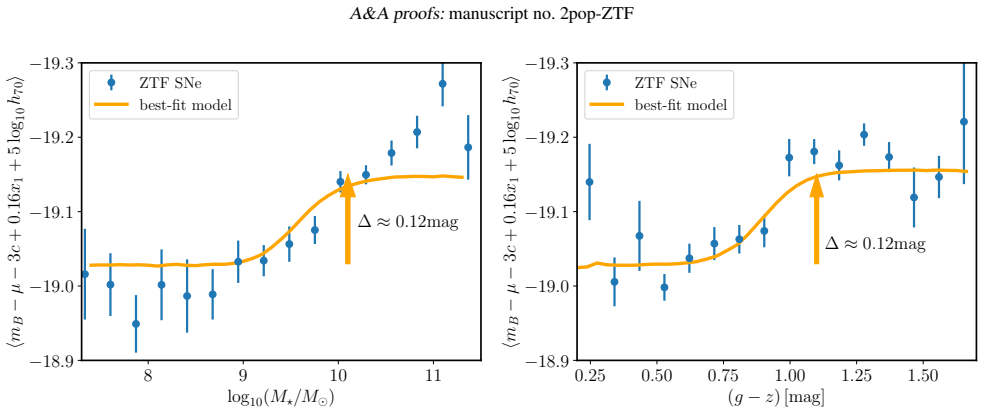
The adopted framework for modelling supernova populations and supernova-host associations makes host-galaxy step corrections redundant. The apparent non-linearity of the supernova magnitude-stretch relation implies a luminosity gap between the supernova populations, with the low-stretch population being 0.14 mag brighter at x1=0, and different slopes. Mean extinction is 3.89 in blue hosts and 3.08 in red hosts.
What carries the argument
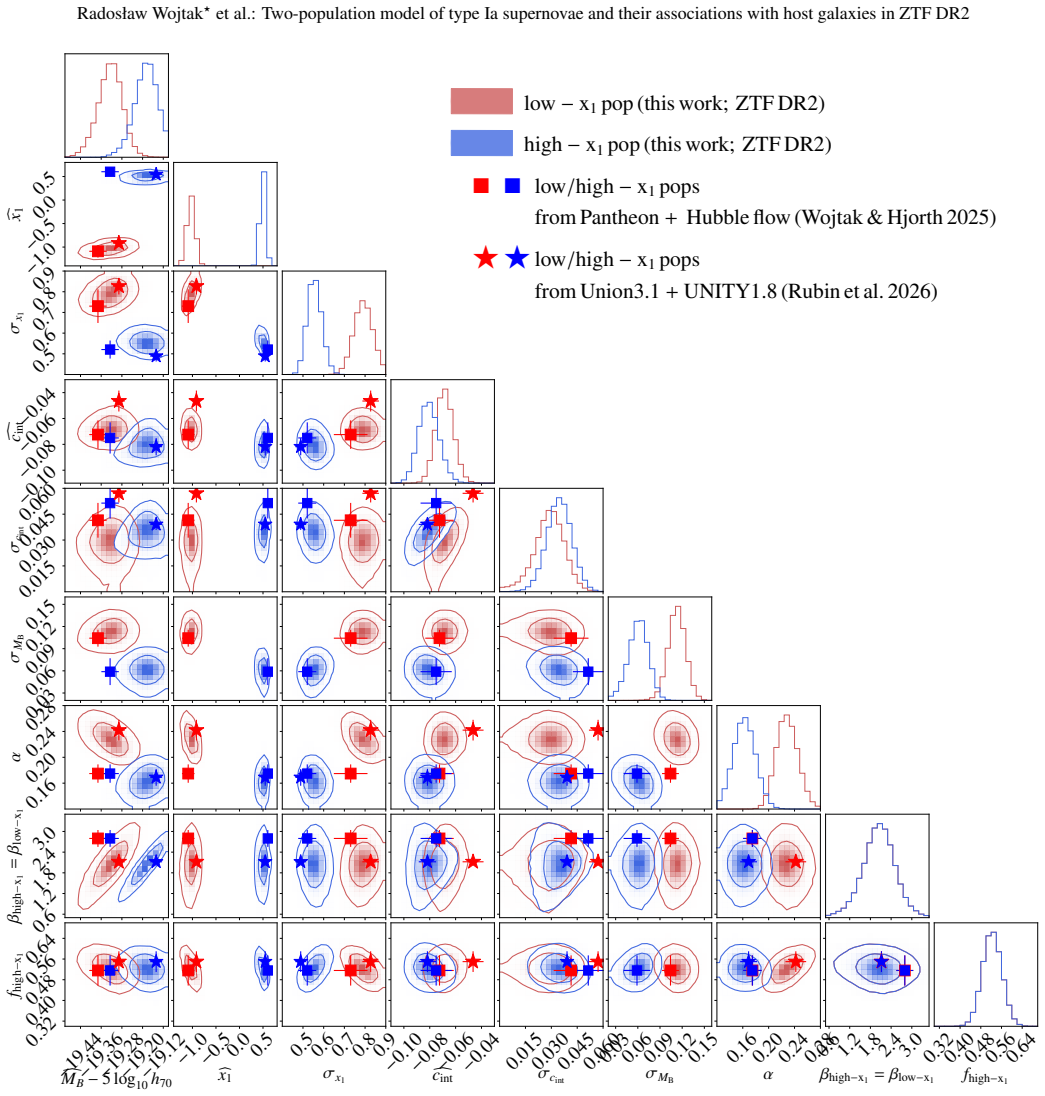
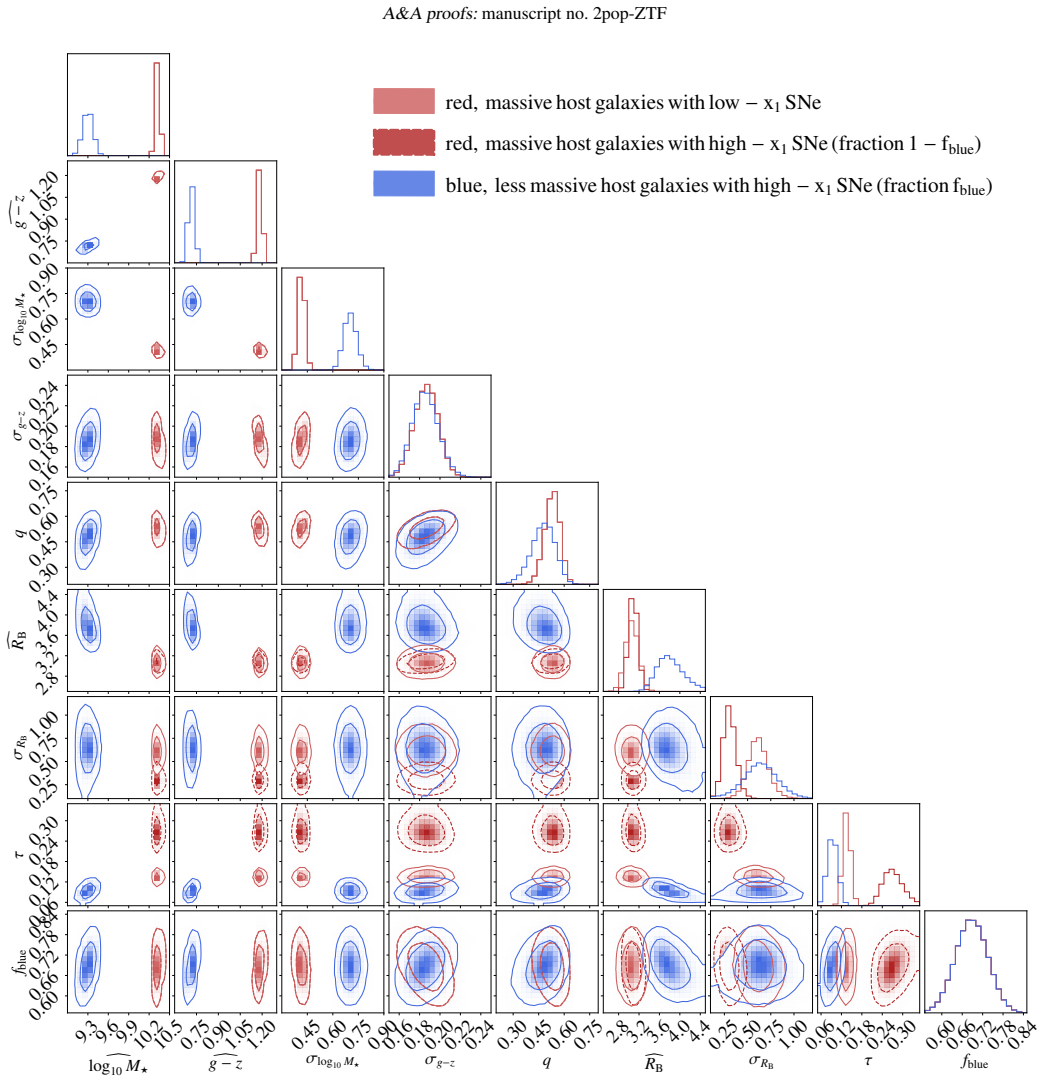
Bayesian hierarchical mixture model of two supernova populations (high-stretch and low-stretch) and two host-galaxy populations (red/massive and blue/less massive), with low-stretch supernovae restricted to red hosts.
If this is right
- Host-galaxy steps in both stellar mass and colour emerge naturally from the way the two supernova populations are distributed across host types.
- The low-stretch population is brighter by 0.14 mag at x1=0 and follows a steeper magnitude-stretch slope than the high-stretch population.
- Mean extinction coefficients differ between host populations, reaching 3.89 in blue hosts versus 3.08 in red hosts.
- No additional parameters for host-galaxy steps are needed once the mixture model and association rules are adopted.
Where Pith is reading between the lines
- The population associations may reflect distinct progenitor channels for high-stretch and low-stretch events.
- Cosmological distance fits could incorporate the mixture model directly rather than applying post-hoc corrections.
- Targeted searches for low-stretch events in blue hosts would provide a direct test of the model's structural assumption.
Load-bearing premise
Low-stretch supernovae are assumed to be exclusively associated with the red/massive host-galaxy population.
What would settle it
A significant fraction of low-stretch type Ia supernovae found in blue or low-mass host galaxies would falsify the assumption about population associations.
Figures
read the original abstract
We constrain type Ia supernova intrinsic properties, extinction, and probabilistic supernova-host associations using the volume-limited sample from the Zwicky Transient Factory DR2, the largest selection-free data set of type Ia supernovae to date. We employ Bayesian hierarchical modelling to jointly analyse the distribution of SALT2 light-curve parameters and global host-galaxy properties (stellar mass and rest-frame g-z colour). The adopted model is a mixture of distributions representing two supernova populations corresponding to two distinct modes of the stretch-parameter distribution, and two host-galaxy populations corresponding to two modes in the host-galaxy parameter space (red/massive and blue/less massive). Motivated by observations, high-stretch supernovae are allowed to populate both host-galaxy populations, whereas low-stretch supernovae are assumed to be exclusively associated with the red/massive host-galaxy population. The apparent non-linearity of the supernova magnitude-stretch relation implies a luminosity gap between the supernova populations, with the low-stretch population being Delta MB=0.14+/-0.03 mag brighter at x1=0, and different slopes (Delta alpha=0.064+/-0.023, steeper for the low-stretch population). The mean extinction coefficient is RB=3.89+/-0.29 (consistent with typical Milky Way values) in the blue/less massive host-galaxy population, which contains 68 per cent of high-stretch supernovae, and RB=3.08+/-0.08 in the red/massive host-galaxy population. Host-galaxy step corrections, both in stellar mass and colour, naturally emerge from the way the two supernova populations, characterised by different intrinsic luminosities and exctinctions, are distributed across host-galaxy populations. The adopted framework for modelling supernova populations and supernova-host associations makes host-galaxy step corrections redundant.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript presents a Bayesian hierarchical mixture model applied to the ZTF DR2 volume-limited Type Ia supernova sample. It jointly fits SALT2 light-curve parameters (stretch x1 and color) with host-galaxy stellar mass and g-z colour, positing two SN populations (high- and low-stretch) and two host populations (red/massive and blue/less massive). Low-stretch SNe are fixed to occur exclusively in red/massive hosts while high-stretch SNe occupy both; the fit yields ΔM_B = 0.14 ± 0.03 mag (low-stretch brighter at x1=0), Δα = 0.064 ± 0.023 (steeper for low-stretch), and R_B = 3.89 ± 0.29 (blue hosts) versus 3.08 ± 0.08 (red hosts). The authors conclude that observed mass and colour steps arise naturally from the differing intrinsic properties and host associations, rendering explicit step corrections redundant.
Significance. If the population-association structure is shown to be robust rather than imposed, the result would supply a physically interpretable mechanism for host-galaxy dependencies in SN Ia standardization. The large, selection-free ZTF DR2 dataset and joint hierarchical treatment of light-curve and host observables provide a concrete empirical foundation that could reduce reliance on ad-hoc corrections in cosmological analyses.
major comments (2)
- [Abstract and model description] Abstract and model description: the exclusivity constraint (low-stretch SNe occur only in red/massive hosts) is imposed as a fixed structural rule rather than inferred from the joint posterior. This choice directly controls how the two SN populations are distributed across hosts and is required for the predicted magnitude offsets and the claim that steps emerge naturally; the manuscript does not report a test in which the constraint is relaxed or replaced by a free mixing parameter.
- [Abstract] Abstract: the reported values of ΔM_B, Δα and the two R_B coefficients are conditional on the fixed association rule. No evidence is presented that these parameters (or the redundancy conclusion) remain stable when the low-stretch population is permitted to occupy blue hosts as well.
minor comments (2)
- [Abstract] Abstract: 'exctinctions' is a typographical error.
- The manuscript would benefit from an explicit figure showing the stretch-magnitude relation for the two populations to illustrate the claimed luminosity gap and slope difference.
Simulated Author's Rebuttal
We thank the referee for the detailed and constructive report. The two major comments correctly identify that the low-stretch exclusivity rule is a fixed modeling choice motivated by observations rather than a fully free parameter, and that the reported results are conditional on this choice. We address both points below and will revise the manuscript accordingly.
read point-by-point responses
-
Referee: Abstract and model description: the exclusivity constraint (low-stretch SNe occur only in red/massive hosts) is imposed as a fixed structural rule rather than inferred from the joint posterior. This choice directly controls how the two SN populations are distributed across hosts and is required for the predicted magnitude offsets and the claim that steps emerge naturally; the manuscript does not report a test in which the constraint is relaxed or replaced by a free mixing parameter.
Authors: The constraint is imposed on the basis of existing observational evidence that low-stretch SNe Ia are found almost exclusively in massive, red hosts. The hierarchical structure still lets the data determine the population fractions, luminosity gap, slope difference, and extinction coefficients. We acknowledge that no explicit test with a relaxed mixing parameter is presented. In the revision we will add a comparison run in which low-stretch events are allowed in both host populations (with an additional free mixing fraction) and report the resulting parameter shifts and the status of the step-redundancy conclusion. revision: yes
-
Referee: Abstract: the reported values of ΔM_B, Δα and the two R_B coefficients are conditional on the fixed association rule. No evidence is presented that these parameters (or the redundancy conclusion) remain stable when the low-stretch population is permitted to occupy blue hosts as well.
Authors: The quoted values and the redundancy statement are indeed conditional on the adopted association structure. Adding the relaxed-mixing model described above will directly test stability of ΔM_B, Δα, the two R_B values, and whether the host steps continue to emerge without explicit corrections. These results will be included in the revised manuscript and discussed in the text. revision: yes
Circularity Check
No significant circularity; results from data-driven Bayesian fit
full rationale
The paper constructs a Bayesian hierarchical mixture model with two SN populations and two host populations, imposing the low-stretch exclusivity to red/massive hosts as a stated modeling choice motivated by prior observations. All reported quantities (ΔM_B, Δα, R_B values, and the redundancy of step corrections) are obtained as posterior constraints from the joint fit to the ZTF DR2 light-curve and host data. No equation or result reduces by construction to a fitted input, self-citation chain, or renamed ansatz; the derivation remains self-contained against the external sample.
Axiom & Free-Parameter Ledger
free parameters (3)
- population mixture weights and distribution parameters for stretch and host properties
- RB extinction coefficients
- luminosity offset Delta MB and slope difference Delta alpha
axioms (2)
- domain assumption Low-stretch supernovae are exclusively associated with the red/massive host-galaxy population
- domain assumption The data are well described by a two-component mixture in both supernova stretch and host-galaxy parameter space
Reference graph
Works this paper leans on
-
[1]
2015, MNRAS, 453, 3300
Amanullah, R., Johansson, J., Goobar, A., et al. 2015, MNRAS, 453, 3300
2015
-
[2]
& Hjorth, J
Andersen, P. & Hjorth, J. 2018, MNRAS, 480, 68
2018
-
[3]
2023, Phys
Balkenhol, L., Dutcher, D., Spurio Mancini, A., et al. 2023, Phys. Rev. D, 108, 023510
2023
-
[4]
2022, A&A, 657, A22
Briday, M., Rigault, M., Graziani, R., et al. 2022, A&A, 657, A22
2022
-
[5]
& Scolnic, D
Brout, D. & Scolnic, D. 2021, ApJ, 909, 26
2021
-
[6]
J., Lavaux, G., & Hudson, M
Carrick, J., Turnbull, S. J., Lavaux, G., & Hudson, M. J. 2015, MNRAS, 450, 317
2015
-
[7]
2025, MNRAS, 538, 3340
Chung, C., Park, S., Son, J., Cho, H., & Lee, Y .-W. 2025, MNRAS, 538, 3340
2025
-
[8]
2023, ApJ, 959, 94
Chung, C., Yoon, S.-J., Park, S., et al. 2023, ApJ, 959, 94
2023
-
[9]
2016, ApJ, 819, 152
Cikota, A., Deustua, S., & Marleau, F. 2016, ApJ, 819, 152
2016
-
[10]
2023, A&A, 680, A56
Duarte, J., González-Gaitán, S., Mourão, A., et al. 2023, A&A, 680, A56
2023
-
[11]
Duarte, J., González-Gaitán, S., Mourão, A. M., Santos, R. P., & Wojtak, R. 2026, arXiv e-prints, arXiv:2605.23512
Pith/arXiv arXiv 2026
-
[12]
Fitzpatrick, E. L. & Massa, D. 2007, ApJ, 663, 320
2007
-
[13]
W., Lang, D., & Goodman, J
Foreman-Mackey, D., Hogg, D. W., Lang, D., & Goodman, J. 2013, PASP, 125, 306
2013
-
[14]
2025, A&A, 694, A4
Ginolin, M., Rigault, M., Copin, Y ., et al. 2025, A&A, 694, A4
2025
-
[15]
2024, arXiv e-prints, arXiv:2405.20965 González-Gaitán, S., de Jaeger, T., Galbany, L., et al
Ginolin, M., Rigault, M., Smith, M., et al. 2024, arXiv e-prints, arXiv:2405.20965 González-Gaitán, S., de Jaeger, T., Galbany, L., et al. 2021, MNRAS, 508, 4656 González-Gaitán, S., Gutiérrez, C. P., Duarte, J., et al. 2026, arXiv e-prints, arXiv:2602.10084
arXiv 2024
- [16]
-
[17]
2007, A&A, 466, 11
Guy, J., Astier, P., Baumont, S., et al. 2007, A&A, 466, 11
2007
-
[18]
Hallgren, L., Wojtak, R., Hjorth, J., & Steinhardt, C. L. 2025, arXiv e-prints, arXiv:2505.22216
arXiv 2025
-
[19]
Kang, Y ., Lee, Y .-W., Kim, Y .-L., Chung, C., & Ree, C. H. 2020, ApJ, 889, 8
2020
-
[20]
L., Hicken, M., Burke, D
Kelly, P. L., Hicken, M., Burke, D. L., Mandel, K. S., & Kirshner, R. P. 2010, ApJ, 715, 743
2010
-
[21]
2013, ApJ, 764, 48
Kessler, R., Guy, J., Marriner, J., et al. 2013, ApJ, 764, 48
2013
-
[22]
2022, MNRAS, 517, 2697
Lee, Y .-W., Chung, C., Demarque, P., et al. 2022, MNRAS, 517, 2697
2022
-
[23]
2013, MNRAS, 436, 222
Maguire, K., Sullivan, M., Patat, F., et al. 2013, MNRAS, 436, 222
2013
-
[24]
S., Thorp, S., Narayan, G., Friedman, A
Mandel, K. S., Thorp, S., Narayan, G., Friedman, A. S., & Avelino, A. 2022, MNRAS, 510, 3939
2022
-
[25]
2006, MNRAS, 370, 773
Mannucci, F., Della Valle, M., & Panagia, N. 2006, MNRAS, 370, 773
2006
-
[26]
2014, ARA&A, 52, 107
Maoz, D., Mannucci, F., & Nelemans, G. 2014, ARA&A, 52, 107
2014
-
[27]
2021, A&A, 649, A74 Padilla Gonzalez, E., Joshi, B
Nicolas, N., Rigault, M., Copin, Y ., et al. 2021, A&A, 649, A74 Padilla Gonzalez, E., Joshi, B. A., Strolger, L. G., et al. 2026, ApJ, 1001, 95
2021
-
[28]
2026, arXiv e-prints, arXiv:2605.12596
Park, S., Lee, Y .-W., Chung, C., et al. 2026, arXiv e-prints, arXiv:2605.12596
Pith/arXiv arXiv 2026
-
[29]
1999, ApJ, 517, 565
Perlmutter, S., Aldering, G., Goldhaber, G., et al. 1999, ApJ, 517, 565
1999
-
[30]
M., Lira, P., Suntzeff, N
Phillips, M. M., Lira, P., Suntzeff, N. B., et al. 1999, AJ, 118, 1766 Planck Collaboration, Aghanim, N., Akrami, Y ., et al. 2020a, A&A, 641, A1 Planck Collaboration, Aghanim, N., Akrami, Y ., et al. 2020b, A&A, 641, A6
1999
-
[31]
2023, ApJ, 945, 84
Popovic, B., Brout, D., Kessler, R., & Scolnic, D. 2023, ApJ, 945, 84
2023
-
[32]
V ., Novinskaya, A
Pruzhinskaya, M. V ., Novinskaya, A. K., Pauna, N., & Rosnet, P. 2020, MNRAS, 499, 5121
2020
-
[33]
J., Wiseman, P., & Sullivan, M
Ramaiya, S., Vincenzi, M., Jarvis, M. J., Wiseman, P., & Sullivan, M. 2025, MNRAS, 543, 2180
2025
-
[34]
G., Filippenko, A
Riess, A. G., Filippenko, A. V ., Challis, P., et al. 1998, AJ, 116, 1009
1998
-
[35]
G., Yuan, W., Macri, L
Riess, A. G., Yuan, W., Macri, L. M., et al. 2022, ApJ, 934, L7
2022
-
[36]
2020, A&A, 644, A176
Rigault, M., Brinnel, V ., Aldering, G., et al. 2020, A&A, 644, A176
2020
-
[37]
2013, A&A, 560, A66
Rigault, M., Copin, Y ., Aldering, G., et al. 2013, A&A, 560, A66
2013
-
[38]
2025, A&A, 694, A1
Rigault, M., Smith, M., Goobar, A., et al. 2025, A&A, 694, A1
2025
-
[39]
A., Riess, A
Rodney, S. A., Riess, A. G., Strolger, L.-G., et al. 2014, AJ, 148, 13
2014
-
[40]
2026, arXiv e-prints, arXiv:2605.14437
Rojas-Bravo, C., Sun, N.-C., Smith, M., et al. 2026, arXiv e-prints, arXiv:2605.14437
Pith/arXiv arXiv 2026
-
[41]
2018, A&A, 615, A68
Roman, M., Hardin, D., Betoule, M., et al. 2018, A&A, 615, A68
2018
-
[42]
M., Popovic, B., Scolnic, D., & Brout, D
Rose, B. M., Popovic, B., Scolnic, D., & Brout, D. 2022, MNRAS, 516, 4822
2022
-
[43]
2026, arXiv e-prints, arXiv:2601.19854
Rubin, D., Hoyt, T., Aldering, G., & Perlmutter, S. 2026, arXiv e-prints, arXiv:2601.19854
arXiv 2026
-
[44]
Salim, S., Boquien, M., & Lee, J. C. 2018, ApJ, 859, 11
2018
-
[45]
& Bildsten, L
Scannapieco, E. & Bildsten, L. 2005, ApJ, 629, L85
2005
-
[46]
F., Meisner, A
Schlafly, E. F., Meisner, A. M., Stutz, A. M., et al. 2016, ApJ, 821, 78
2016
-
[47]
M., Jones, D
Scolnic, D. M., Jones, D. O., Rest, A., et al. 2018, ApJ, 859, 101
2018
-
[48]
2025, A&A, 694, A14
Senzel, R., Maguire, K., Burgaz, U., et al. 2025, A&A, 694, A14
2025
-
[49]
A., et al
Sullivan, M., Conley, A., Howell, D. A., et al. 2010, MNRAS, 406, 782
2010
-
[50]
J., et al
Sullivan, M., Le Borgne, D., Pritchet, C. J., et al. 2006, ApJ, 648, 868
2006
-
[51]
& Mandel, K
Thorp, S. & Mandel, K. S. 2022, MNRAS, 517, 2360
2022
-
[52]
S., Jones, D
Thorp, S., Mandel, K. S., Jones, D. O., Ward, S. M., & Narayan, G. 2021, MN- RAS, 508, 4310
2021
-
[53]
2025, MNRAS, 538, 181
Toy, M., Wiseman, P., Sullivan, M., et al. 2025, MNRAS, 538, 181
2025
-
[54]
Tucker, M. A. 2025, MNRAS, 538, L1
2025
-
[55]
2024, ApJ, 975, 86
Vincenzi, M., Brout, D., Armstrong, P., et al. 2024, ApJ, 975, 86
2024
-
[56]
2022, MNRAS, 515, 4587
Wiseman, P., Vincenzi, M., Sullivan, M., et al. 2022, MNRAS, 515, 4587
2022
-
[57]
& Hjorth, J
Wojtak, R. & Hjorth, J. 2022, MNRAS, 515, 2790
2022
-
[58]
& Hjorth, J
Wojtak, R. & Hjorth, J. 2025, A&A, 702, A176
2025
-
[59]
Wojtak, R., Hjorth, J., & Hjortlund, J. O. 2023, MNRAS, 525, 5187
2023
-
[60]
Yoon, S.-J., Park, I., Zee, W.-B. G., et al. 2026, arXiv e-prints, arXiv:2604.24846 Article number, page 12 of 14 Radosław Wojtak‹ et al.: Two-population model of type Ia supernovae and their associations with host galaxies in ZTF DR2 Appendix A: Supplementary figures 0.0 0.2 0.4 0.6 c 7.5 9.0 10 .5 log 10 (M?/M) 2 1 0 1 x1 0.4 0.8 1.2 1.6 (g z) 0.0 0.2 0...
Pith/arXiv arXiv 2026
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.