Novel Triple-Based Problems for the Construction of Phylogenetic Networks via Least Common Ancestors
Pith reviewed 2026-06-25 21:33 UTC · model grok-4.3
The pith
Phylogenetic networks can realize consistent sets of rooted and anchored triples under an LCA interpretation, with the realizing network constructible in polynomial time.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
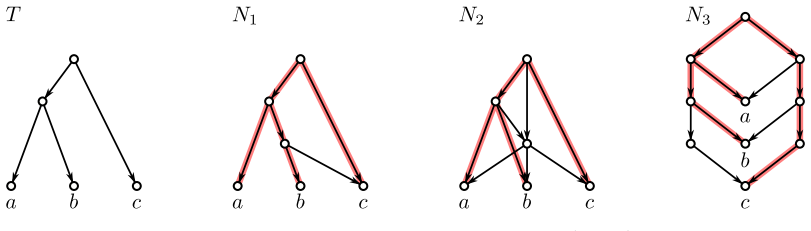
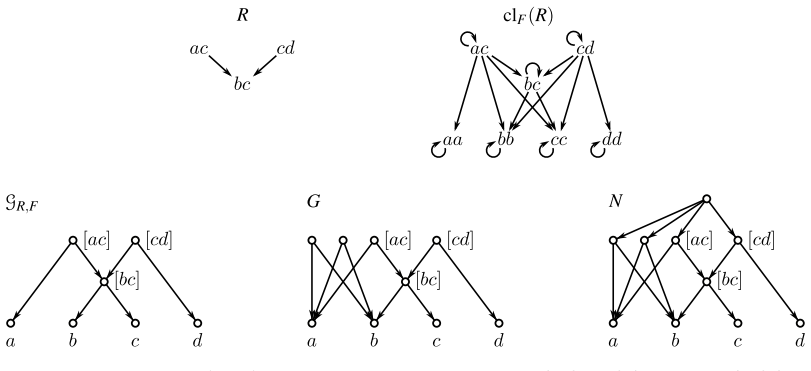
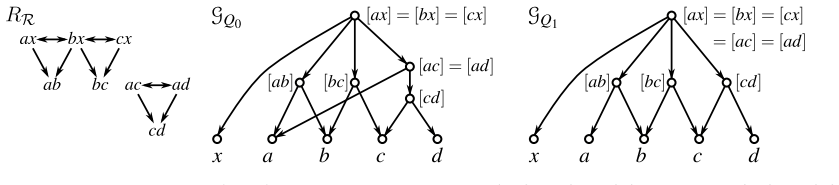
By translating the consistency questions for ordinary and anchored triples into realization problems for required and forbidden LCA-constraints, we show that all resulting problems can be solved in polynomial time. Moreover, whenever a solution exists, a suitable realizing DAG and phylogenetic network can be constructed within the same time bound.
What carries the argument
The reduction of triple consistency questions (with and without forbidden triples) to realization problems for required and forbidden LCA-constraints.
If this is right
- All variants of the consistency problems for triples and anchored triples are solvable in polynomial time.
- A realizing directed acyclic graph exists and can be constructed in polynomial time when the constraints are consistent.
- A phylogenetic network realizing the constraints can be constructed in polynomial time when a solution exists.
- The same polynomial-time bound holds whether or not forbidden triples are present.
Where Pith is reading between the lines
- The approach may extend to consistency problems involving other local constraints on ancestral relationships in networks.
- It suggests that local LCA information extracted from sequences is often sufficient to determine global network structure under these definitions.
- Similar reductions might apply to network inference tasks that incorporate additional data types beyond triples.
Load-bearing premise
The ancestor-based interpretation of triples and the anchored relaxation accurately model the biological information inferable from genomic sequences about relative evolutionary proximity.
What would settle it
An input set of triples (ordinary or anchored, with or without forbidden triples) for which the polynomial-time algorithm outputs a DAG or network that fails to display the required LCA relationships as defined in the paper.
Figures

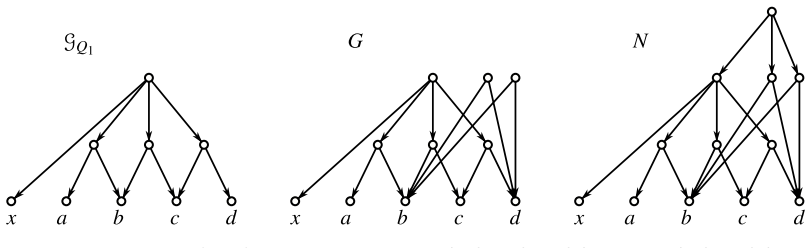
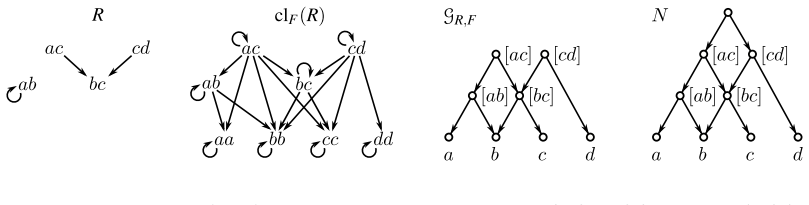
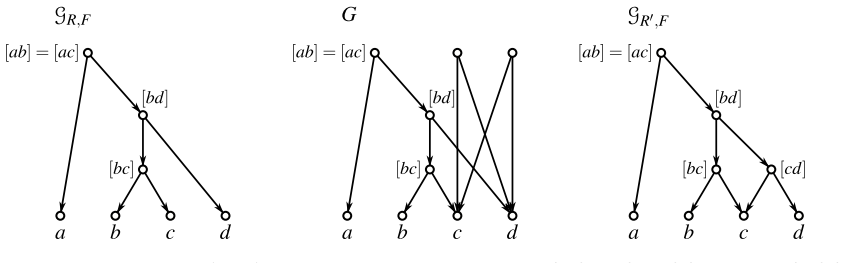
read the original abstract
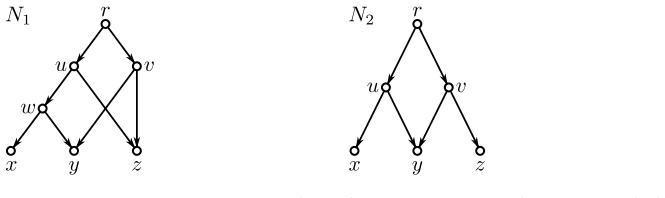
Evolutionary histories are often represented by rooted phylogenetic networks, whose leaves correspond to extant taxa and whose internal vertices represent ancestral lineages. Since such histories must usually be inferred from incomplete data, in particular from genomic sequences of present-day taxa, one often obtains only local information about relative evolutionary proximity. For instance, sequence data may suggest that two taxa $x$ and $y$ are more closely related to each other than either is to a third taxon $z$. This information is classically encoded by a rooted triple $xy|z$. In this paper, we study rooted triples in phylogenetic networks under an ancestor-based interpretation: $xy|z$ is displayed if the unique least common ancestor (LCA) of $x$ and $y$ lies strictly below the unique LCA of $x$ and $z$, respectively of $y$ and $z$, and the latter two LCAs coincide. We also introduce anchored triples $\underline{x}y|z$, which retain only the asymmetric comparison that the LCA of $x$ and $y$ lies below the LCA of $x$ and $z$. This relaxation is natural in networks, where different pairwise ancestral relationships need not behave as they do in trees. We consider several variants of consistency problems for ordinary and anchored triples, both with and without forbidden triples. Somewhat surprisingly, these ancestor-based consistency questions for triples in phylogenetic networks do not appear to have been addressed before despite their direct biological interpretation and the fact that such constraints can be inferred naturally from genomic sequence data. By translating these questions into realization problems for required and forbidden LCA-constraints, we show that all resulting problems can be solved in polynomial time. Moreover, whenever a solution exists, a suitable realizing DAG and phylogenetic network can be constructed within the same time bound.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper defines an ancestor-based display condition for rooted triples xy|z (and anchored variants) in phylogenetic networks, requiring unique LCAs with the LCA of x,y strictly below the coinciding LCAs of the other pairs. It studies consistency problems for sets of such triples (with and without forbidden triples) and claims that all variants are solvable in polynomial time by translation to required/forbidden LCA-constraint realization problems on DAGs; moreover, a realizing phylogenetic network can be constructed in the same time bound whenever a solution exists.
Significance. If the reduction is valid, the results supply the first polynomial-time algorithms and constructions for these local, biologically interpretable constraints on networks (as opposed to trees), addressing a previously unstudied class of problems with direct applicability to inference from genomic data.
major comments (1)
- [Abstract] Abstract (and the definition of triple display): the ancestor-based condition explicitly requires that LCAs are unique for the relevant leaf pairs. However, the claimed polynomial-time reduction to general LCA-constraint realization on DAGs does not appear to guarantee uniqueness of LCAs in the output networks (especially when reticulations are present). This raises a question about whether the equivalence between triple consistency and constraint satisfaction holds in both directions, which is load-bearing for the polynomial-time claim.
Simulated Author's Rebuttal
We thank the referee for their careful reading of the manuscript and for highlighting this important point about the uniqueness of LCAs. We address the comment below.
read point-by-point responses
-
Referee: [Abstract] Abstract (and the definition of triple display): the ancestor-based condition explicitly requires that LCAs are unique for the relevant leaf pairs. However, the claimed polynomial-time reduction to general LCA-constraint realization on DAGs does not appear to guarantee uniqueness of LCAs in the output networks (especially when reticulations are present). This raises a question about whether the equivalence between triple consistency and constraint satisfaction holds in both directions, which is load-bearing for the polynomial-time claim.
Authors: The definition in the manuscript does require unique LCAs, as the referee correctly notes. Our reduction translates the ancestor-based (and anchored) triple conditions into a collection of required and forbidden LCA constraints on a DAG; these constraints are formulated so that any realizing DAG must contain, for each constrained triple, a single vertex that serves as the unique LCA satisfying the stated ancestor relations (equality of the two higher LCAs and strict descent of the lower one). The known polynomial-time algorithm for LCA-constraint realization constructs precisely such a minimal DAG, in which the constrained pairs have unique LCAs by construction of the partial order on ancestor vertices. Reticulations are permitted but do not create additional minimal common ancestors for the constrained pairs because the equality and descent constraints force a unique meeting point. We will revise the manuscript to add an explicit paragraph clarifying this direction of the equivalence and confirming that the output networks satisfy the uniqueness requirement in the original definition. revision: yes
Circularity Check
No circularity; algorithmic translation and poly-time claims are independent of inputs
full rationale
The paper defines ancestor-based and anchored triple display via unique-LCA predicates, translates consistency questions into LCA-constraint realization problems, and asserts polynomial-time solvability plus explicit DAG/network construction. No fitted parameters, no self-referential definitions (the display predicate is stated directly, not derived from the output networks), and no load-bearing self-citations. The derivation consists of standard reduction to a constraint-realization task whose complexity is established separately; the result does not reduce to its own inputs by construction. The skeptic concern about uniqueness of LCAs in the output networks is a potential soundness issue for the equivalence, not a circularity in the derivation chain itself.
Axiom & Free-Parameter Ledger
axioms (1)
- standard math Least common ancestors exist and are unique for pairs of leaves in the considered rooted phylogenetic networks.
Reference graph
Works this paper leans on
-
[1]
SIAM Journal on Computing 10(3):405–421, DOI 10.1137/0210030 29
Aho A V, Sagiv Y, Szymanski TG, Ullman JD (1981) Inferring a tree from lowest common ancestors with an application to the optimization of relational expressions. SIAM Journal on Computing 10(3):405–421, DOI 10.1137/0210030 29
-
[2]
Springer Monographs in Mathematics, Springer
Bang-Jensen J, Gutin GZ (2009) Digraphs: Theory, Algorithms and Applications, 2nd edn. Springer Monographs in Mathematics, Springer
2009
-
[3]
theory and methods in phylogenetic analysis
Bryant D (1997) Building trees, hunting for trees, and comparing trees. theory and methods in phylogenetic analysis. PhD thesis, University of Canterbury, DOI 10.26021/2479
-
[4]
Bryant D, Steel M (1995) Extension operations on sets of leaf-labeled trees. Adv Appl Math 16(4):425–453, DOI 10.1006/aama.1995.1020
-
[5]
Discrete Applied Mathematics 127(2):241–269, DOI 10.1016/S0166-218X(02) 00209-3
Caspard N, Monjardet B (2003) The lattices of closure systems, closure operators, and implicational systems on a finite set: a survey. Discrete Applied Mathematics 127(2):241–269, DOI 10.1016/S0166-218X(02) 00209-3
-
[6]
MIT press
Cormen TH, Leiserson CE, Rivest RL, Stein C (2022) Introduction to Algorithms. MIT press
2022
-
[7]
URLhttps://arxiv.org/abs/2605.03827,2605.03827
Ebert PA, Hellmuth M (2026) Inferring phylogenetic networks from required and forbidden LCA- constraints. URLhttps://arxiv.org/abs/2605.03827,2605.03827
Pith/arXiv arXiv 2026
-
[8]
Journal of Mathematical Biology 74(7):1729–1751, DOI 10.1007/s00285-016-1068-3
Gambette P, Huber KT, Kelk S (2017) On the challenge of reconstructing level-1 phylogenetic networks from triplets and clusters. Journal of Mathematical Biology 74(7):1729–1751, DOI 10.1007/s00285-016-1068-3
-
[9]
Journal of Mathematical Biology 78(7):2015– 2057, DOI 10.1007/s00285-019-01332-9
Geiß M, Ch ´avez E, Gonz ´alez Laffitte M, L ´opez S ´anchez A, Stadler BMR, Valdivia DI, Hellmuth M, Hern´andez Rosales M, Stadler PF (2019) Best match graphs. Journal of Mathematical Biology 78(7):2015– 2057, DOI 10.1007/s00285-019-01332-9
-
[10]
Journal of Mathematical Biology 80(3):865–953, DOI 10.1007/s00285-019-01444-2
Geiß M, Stadler PF, Hellmuth M (2020) Reciprocal best match graphs. Journal of Mathematical Biology 80(3):865–953, DOI 10.1007/s00285-019-01444-2
-
[11]
Gr¨ unewald S, Spillner A, Bastkowski S, Bogershausen A, Moulton V (2013) Superq: Computing supernet- works from quartets. IEEE/ACM Transactions on Computational Biology and Bioinformatics 10(1):151– 160, DOI 10.1109/TCBB.2013.8
-
[12]
Journal of Bioinformatics and Computational Biology 04(01):59–74, DOI 10.1142/S0219720006001709
He YJ, Huynh TND, Jansson J, Sung WK (2006) Inferring phylogenetic relationships avoiding for- bidden rooted triplets. Journal of Bioinformatics and Computational Biology 04(01):59–74, DOI 10.1142/S0219720006001709
-
[13]
URLhttps://arxiv.org/abs/2606.16963,2606.16963
Hellmuth M, Lindeberg A, Moulton V (2026) Encoding phylogenetic networks with least common ancestor constraints. URLhttps://arxiv.org/abs/2606.16963,2606.16963
arXiv 2026
-
[14]
Algorithmica 66(3):714–738, DOI 10.1007/s00453-012-9659-x
Huber KT, Moulton V (2013) Encoding and constructing 1-nested phylogenetic networks with trinets. Algorithmica 66(3):714–738, DOI 10.1007/s00453-012-9659-x
-
[15]
Huber KT, van Iersel L, Kelk S, Suchecki R (2011) A practical algorithm for reconstructing level-1 phylogenetic networks. IEEE/ACM Transactions on Computational Biology and Bioinformatics 8(3):635– 649, DOI 10.1109/TCBB.2010.17
-
[16]
Algorithmica 77(1):173–200, DOI 10.1007/s00453-015-0069-8
Huber KT, van Iersel L, Moulton V, Scornavacca C, Wu T (2017) Reconstructing phylogenetic level-1 net- works from nondense binet and trinet sets. Algorithmica 77(1):173–200, DOI 10.1007/s00453-015-0069-8
-
[17]
Huson D, Dezulian T, Klopper T, Steel M (2004) Phylogenetic super-networks from partial trees. IEEE/ACM Transactions on Computational Biology and Bioinformatics 1(4):151–158, DOI 10.1109/TCBB.2004.44
-
[18]
Genome Biol Evol 3:23–35, DOI 10.1093/gbe/evq077
Huson DH, Scornavacca C (2011) A survey of combinatorial methods for phylogenetic networks. Genome Biol Evol 3:23–35, DOI 10.1093/gbe/evq077
-
[19]
Huson DH, Dezulian T, Kl ¨opper T, Steel MA (2004) Phylogenetic super-networks from partial trees. In: Jonassen I, Kim J (eds) Algorithms in Bioinformatics, Springer Berlin Heidelberg, Berlin, Heidelberg, pp 388–399, DOI 10.1007/978-3-540-30219-3 33
-
[20]
Cambridge University Press
Huson DH, Rupp R, Scornavacca C (2011) Phylogenetic Networks: Concepts, Algorithms and Applications. Cambridge University Press
2011
-
[21]
Algo- rithmica 60(2):207–235, DOI 10.1007/s00453-009-9333-0 30
van Iersel L, Kelk S (2011) Constructing the simplest possible phylogenetic network from triplets. Algo- rithmica 60(2):207–235, DOI 10.1007/s00453-009-9333-0 30
-
[22]
Journal of Mathematical Biology 68(7):1707–1729, DOI 10.1007/s00285-013-0683-5
van Iersel L, Moulton V (2014) Trinets encode tree-child and level-2 phylogenetic networks. Journal of Mathematical Biology 68(7):1707–1729, DOI 10.1007/s00285-013-0683-5
-
[23]
van Iersel L, Keijsper J, Kelk S, Stougie L, Hagen F, Boekhout T (2009) Constructing level-2 phylogenetic networks from triplets. IEEE/ACM Transactions on Computational Biology and Bioinformatics 6(4):667– 681, DOI 10.1109/TCBB.2009.22
-
[24]
Journal of Bioinformatics and Computational Biology 07(04):597–623, DOI 10.1142/S0219720009004308
van Iersel L, Kelk S, Mnich M (2009) Uniqueness, intractability and exact algorithms: Reflections on level-k phylogenetic networks. Journal of Bioinformatics and Computational Biology 07(04):597–623, DOI 10.1142/S0219720009004308
-
[25]
Information Processing Letters 178:106300, DOI 10.1016/j.ipl.2022.106300
van Iersel L, Kole S, Moulton V, Nipius L (2022) An algorithm for reconstructing level-2 phylogenetic networks from trinets. Information Processing Letters 178:106300, DOI 10.1016/j.ipl.2022.106300
-
[26]
Jansson J, Sung WK (2006) Inferring a level-1 phylogenetic network from a dense set of rooted triplets. Theoretical Computer Science 363(1):60–68, DOI 10.1016/j.tcs.2006.06.022, computing and Combina- torics
-
[27]
SIAM Journal on Computing 35(5):1098–1121, DOI 10.1137/S0097539704446529
Jansson J, Nguyen NB, Sung WK (2006) Algorithms for combining rooted triplets into a galled phylogenetic network. SIAM Journal on Computing 35(5):1098–1121, DOI 10.1137/S0097539704446529
-
[28]
Kowaluk M, Lingas A (2007) Unique lowest common ancestors in dags are almost as easy as matrix multiplication. In: Arge L, Hoffmann M, Welzl E (eds) Algorithms – ESA 2007, Springer Berlin Heidelberg, pp 265–274, DOI 10.1007/978-3-540-75520-3 25
-
[29]
Bull Math Biol 87(3):44, DOI 10.1007/s11538-025-01419-z
Lindeberg A, Hellmuth M (2025) Simplifying and characterizing DAGs and phylogenetic networks via least common ancestor constraints. Bull Math Biol 87(3):44, DOI 10.1007/s11538-025-01419-z
-
[30]
Displaying trees across two phylogenetic networks
Lindeberg A, Alfonsson A, Moulton V, Scholz GE, Hellmuth M (2026) Inferring DAGs and phylogenetic networks from least common ancestors. Theoretical Computer Science 1082:116114, DOI 10.1016/j.tcs. 2026.116114
-
[31]
Oxford University Press
Matou ˇsek J, Neˇsetˇril J (2009) Invitation to discrete mathematics. Oxford University Press
2009
-
[32]
Discrete Applied Mathematics 98(3):227–235, DOI 10.1016/S0166-218X(99)00160-2
Ng MP, Steel M, Wormald NC (2000) The difficulty of constructing a leaf-labelled tree including or avoiding given subtrees. Discrete Applied Mathematics 98(3):227–235, DOI 10.1016/S0166-218X(99)00160-2
-
[33]
PLOS ONE 9(9):1–1, DOI 10.1371/journal.pone.0106531
Poormohammadi H, Eslahchi C, Tusserkani R (2014) Tripnet: A method for constructing rooted phyloge- netic networks from rooted triplets. PLOS ONE 9(9):1–1, DOI 10.1371/journal.pone.0106531
-
[34]
Theoretical Computer Science 865:63–84, DOI 10.1016/j.tcs.2021.02.037
Schaller D, Stadler PF, Hellmuth M (2021) Complexity of modification problems for best match graphs. Theoretical Computer Science 865:63–84, DOI 10.1016/j.tcs.2021.02.037
-
[35]
Birkh¨auser Boston, DOI 10.1007/978-1-4612-0053-6
Schr ¨oder BSW (2003) Ordered Sets. Birkh¨auser Boston, DOI 10.1007/978-1-4612-0053-6
-
[36]
European Journal of Combinatorics 70:384–407, DOI 10.1016/j.ejc.2018.02.013
Seemann CR, Hellmuth M (2018) The matroid structure of representative triple sets and triple-closure computation. European Journal of Combinatorics 70:384–407, DOI 10.1016/j.ejc.2018.02.013
-
[37]
Journal of Mathematical Biology 83(3):28, DOI 10.1007/s00285-021-01654-7
Semple C, Toft G (2021) Trinets encode orchard phylogenetic networks. Journal of Mathematical Biology 83(3):28, DOI 10.1007/s00285-021-01654-7
-
[38]
Willson SJ (2011) Regular networks can be uniquely constructed from their trees. IEEE/ACM Transactions on Computational Biology and Bioinformatics 8(3):785–796, DOI 10.1109/TCBB.2010.69 31
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.