Nullclines, Subnullclines and the Asymptotic and Transient Attractors in Eco-Evolutionary Dynamics
Pith reviewed 2026-06-27 20:14 UTC · model grok-4.3
The pith
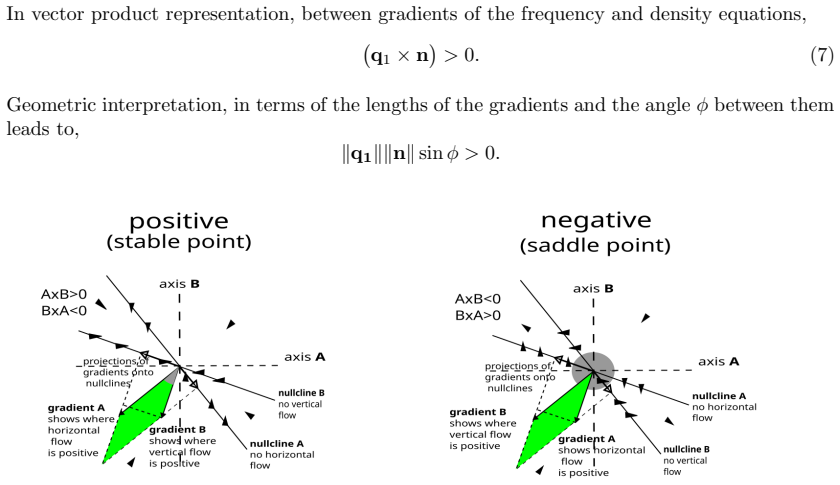
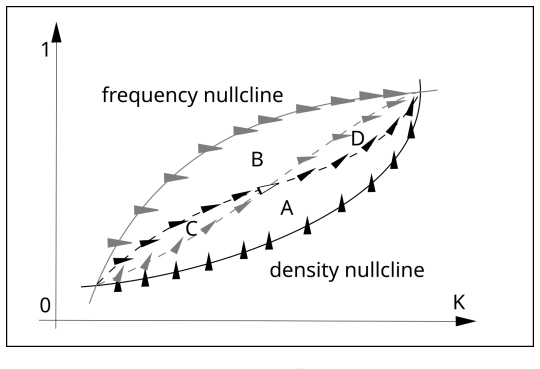
In the Hawk-Dove game with separate mortality and fertility payoffs, frequency and density nullclines intersect at equilibria linked by heteroclinic orbits that reveal subnullclines as intermediate manifolds attracting trajectories before c
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The stable and unstable equilibria determined by intersections of frequency and density nullclines are connected by heteroclinic orbits that attract nearby trajectories; the resulting bundles expose subnullclines lying between the frequency and density nullclines before trajectories converge to the stable rest point. Adding periodic environmental seasonality produces complex cycling in which the subnullclines serve as resilience thresholds that block propagation of external perturbations.
What carries the argument
Heteroclinic orbits connecting equilibria at nullcline intersections, which generate subnullclines as manifolds that organize transient approach to stable rest points.
If this is right
- Trajectories starting near unstable equilibria are funneled along heteroclinic connections into subnullcline regions before reaching the stable point.
- Subnullclines act as organized transient structures that organize the approach to long-term rest points in the demographic Hawk-Dove model.
- Periodic seasonal mortality generates cycling whose amplitude is constrained by the locations of the subnullclines.
- Subnullclines function as barriers that determine how far an external perturbation can spread before the system returns to its cycle.
Where Pith is reading between the lines
- The same nullcline-plus-subnullcline structure may appear in other matrix games once mortality and fertility payoffs are separated by the same demographic rules.
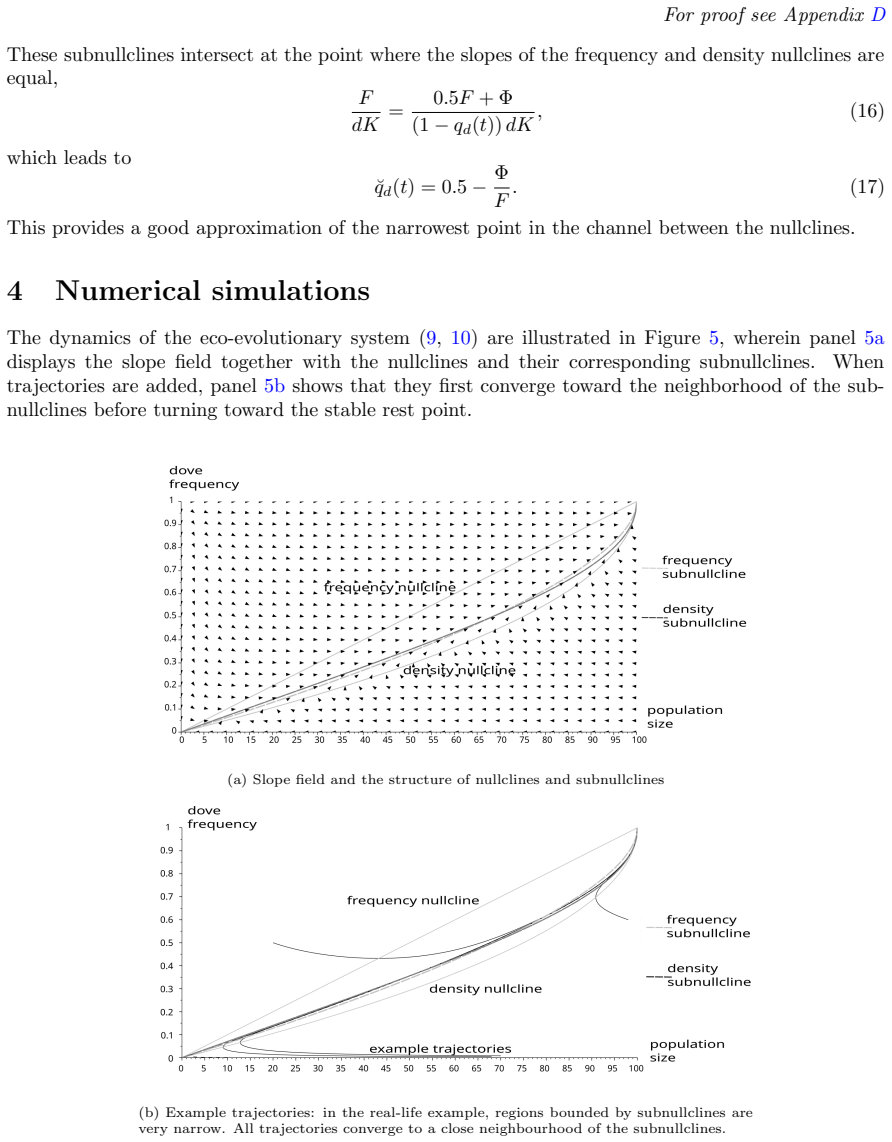
- Subnullclines could be used to define quantitative resistance thresholds in models of seasonal population management or conservation.
- If subnullclines persist under stochastic perturbations, they would provide a geometric explanation for why some eco-evolutionary systems recover quickly from shocks while others do not.
Load-bearing premise
Mortality payoff is determined solely by the opponent's strategy while fertility payoff is modified by density-dependent juvenile recruitment survival.
What would settle it
Numerical integration of the system equations that produces trajectories crossing directly from one equilibrium to another without first aligning along manifolds between the frequency and density nullclines.
Figures











read the original abstract
In the demographic framework, mortality payoff function describes the cost of an interaction and fertility payoff function describes its reward. So while mortality cost depends on opponent's strategy, fertility reward can be affected by the density-dependent juvenile recruitment survival. This motivates an analysis of the eco-evolutionary dynamics of the classical Hawk-Dove game. It is shown that the stable and unstable equilibria (determined by the intersections of frequency and density nullclines) are connected by heteroclinic orbits, which attract nearby trajectories. The resulting bundle of trajectories leads to the discovery of the so-called subnullcines (manifolds placed between frequency and density nullcline) before they converge to the stable rest point. The initial isolated system is then extended by adding environmental seasonality (periodic background mortality), which acts as an external factor. This leads to complex cycling behavior and the subnullclines act as barriers to the propagation of the perturbation (resilience/resistance threshold). Thus, in a way, this paper completes, yet extends, previous works on the eco-evolutionary dynamics of games with demographic payoffs.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper analyzes the eco-evolutionary dynamics of the classical Hawk-Dove game in a demographic payoff framework, where mortality costs depend only on the opponent's strategy while fertility rewards are modulated by density-dependent juvenile recruitment survival. It claims that the stable and unstable equilibria (at intersections of frequency and density nullclines) are connected by heteroclinic orbits attracting nearby trajectories; the resulting bundle of trajectories reveals subnullclines (manifolds between the nullclines) as transient structures before convergence to the stable rest point. The model is then extended with periodic environmental seasonality, producing complex cycling in which subnullclines function as barriers to perturbation propagation, thereby extending prior work on demographic payoffs.
Significance. If the derivations are rigorous, the work supplies a geometric phase-plane account of transient attractors and resilience thresholds in eco-evolutionary systems. The separation of mortality and fertility payoffs is a modeling choice that yields distinct nullclines; once adopted, the heteroclinic structure follows from standard planar dynamical-systems reasoning. The introduction of subnullclines as intermediate manifolds and their role under seasonality could provide useful analytic tools for other frequency-density models, provided the definitions are shown to be intrinsic to the vector field rather than trajectory-dependent.
major comments (2)
- [Abstract / Model definition] Abstract and model section: the central claim that subnullclines are discovered from the bundle of heteroclinic orbits requires an explicit, equation-level definition (e.g., as level sets of a Lyapunov-like function or invariant manifolds of the planar system) rather than a descriptive placement “between frequency and density nullclines.” Without this, it remains unclear whether the objects are derived from the vector field or identified post-hoc from numerical trajectories.
- [Model definition / Payoff functions] The separation of payoffs (mortality depending solely on opponent strategy, fertility on density-dependent survival) is presented as the key modeling choice that produces distinct nullclines. The manuscript should state the explicit payoff functions and the resulting ODE system (frequency and density equations) so that the intersections and the claimed heteroclinic connections can be verified directly; the current description leaves open whether the geometry follows by construction or requires additional assumptions.
minor comments (1)
- [Abstract] The abstract contains no equations, derivations, or numerical verification; adding a compact statement of the governing ODEs and the definition of subnullclines would improve readability and allow immediate assessment of the claims.
Simulated Author's Rebuttal
We thank the referee for their careful reading and constructive comments on the manuscript. We address each major comment below and have revised the paper accordingly to improve clarity and rigor.
read point-by-point responses
-
Referee: [Abstract / Model definition] Abstract and model section: the central claim that subnullclines are discovered from the bundle of heteroclinic orbits requires an explicit, equation-level definition (e.g., as level sets of a Lyapunov-like function or invariant manifolds of the planar system) rather than a descriptive placement “between frequency and density nullclines.” Without this, it remains unclear whether the objects are derived from the vector field or identified post-hoc from numerical trajectories.
Authors: We agree that an explicit, equation-level definition is required to establish that subnullclines are intrinsic to the vector field. In the revised manuscript we define them as the unique invariant manifolds lying strictly between the frequency and density nullclines that are transversely attracting; this is shown by constructing a Lyapunov-like function V whose orbital derivative is strictly negative in a neighborhood of each manifold, confirming they are not trajectory-dependent artifacts. The definition and its verification have been added to the model section. revision: yes
-
Referee: [Model definition / Payoff functions] The separation of payoffs (mortality depending solely on opponent strategy, fertility on density-dependent survival) is presented as the key modeling choice that produces distinct nullclines. The manuscript should state the explicit payoff functions and the resulting ODE system (frequency and density equations) so that the intersections and the claimed heteroclinic connections can be verified directly; the current description leaves open whether the geometry follows by construction or requires additional assumptions.
Authors: We accept that the explicit payoff functions and ODE system were not stated with sufficient precision. The revised manuscript now gives the mortality payoff M(s') depending only on the opponent’s strategy and the fertility payoff F(s,s',N) incorporating density-dependent juvenile survival. The resulting planar system is written explicitly as dp/dt = p(1-p)[payoff difference] and dN/dt = N[birth rate(N) – death rate], from which the nullclines, their intersections, and the heteroclinic connections follow directly by standard phase-plane analysis without further assumptions. revision: yes
Circularity Check
No significant circularity detected
full rationale
The paper's core derivation defines frequency and density nullclines from the demographic payoff structure (mortality depending on opponent strategy, fertility modulated by density-dependent survival), then applies standard planar dynamical-systems results to establish heteroclinic orbits connecting equilibria and the existence of intermediate subnullclines as manifolds. These steps follow directly from the chosen model equations without reducing to fitted parameters, self-referential definitions, or load-bearing self-citations that presuppose the target result. The extension to seasonal perturbations and resilience thresholds likewise proceeds from the established phase-plane geometry. The work acknowledges prior papers on demographic payoffs but treats them as background rather than as the sole justification for the new claims, leaving the derivation self-contained against external mathematical benchmarks.
Axiom & Free-Parameter Ledger
axioms (1)
- standard math Standard properties of nullclines, equilibria, and heteroclinic orbits in planar dynamical systems
invented entities (1)
-
subnullclines
no independent evidence
Reference graph
Works this paper leans on
-
[1]
Doebeli, Y
M. Doebeli, Y. Ispolatov, and B. Simon. Towards a mechanistic foundation of evolutionary theory.eLife, 6:e23804, 2017
2017
-
[2]
Argasinski and M
K. Argasinski and M. Broom. Ecological theatre and the evolutionary game: how environmental and demographic factors determine payoffs in evolutionary games.Journal of Mathematical Biology, 67:935–962, 2013
2013
-
[3]
Argasinski and M
K. Argasinski and M. Broom. Interaction rates, vital rates, background fitness and replicator dynamics: how to embed evolutionary game structure into realistic population dynamics.Theory in Biosciences, 137:33–50, 2018
2018
-
[4]
Argasinski and M
K. Argasinski and M. Broom. Evolutionary stability under limited population growth: Eco- evolutionary feedbacks and replicator dynamics.Ecological Complexity, 34:198–212, 2018
2018
-
[5]
Maynard Smith and G
J. Maynard Smith and G. R. Price. The logic of animal conflict.Nature, 246:15, 1973
1973
-
[6]
Maynard Smith.Evolution and the Theory of Games
J. Maynard Smith.Evolution and the Theory of Games. Cambridge University Press, Cambridge, 1982
1982
-
[7]
Hofbauer and K
J. Hofbauer and K. Sigmund.The Theory of Evolution and Dynamical Systems, volume 7 of London Mathematical Society Student Texts. Cambridge University Press, 1988
1988
-
[8]
Hofbauer and K
J. Hofbauer and K. Sigmund.Evolutionary Games and Population Dynamics. Cambridge Uni- versity Press, Cambridge, 1998
1998
-
[9]
Broom and J
M. Broom and J. Rycht´ aˇ r.Game-theoretical Models in Biology. CRC Press, 2022
2022
-
[10]
Friedman and B
D. Friedman and B. Sinervo.Evolutionary Games in Natural, Social, and Virtual Worlds. Oxford University Press, 2016
2016
-
[11]
M. S. Bedi, K. Argasinski, and M. Broom. Eco-evolutionary game dynamics with three strategies: how geometric analysis provides insights into stability.bioRxiv, 2025
2025
-
[12]
Hastings
A. Hastings. Transients: the key to long-term ecological understanding?Trends in Ecology & Evolution, 19(1):39–45, 2004
2004
-
[13]
Hastings, K
A. Hastings, K. C. Abbott, K. Cuddington, T. Francis, G. Gellner, Y. C. Lai, and M. L. Zeeman. Transient phenomena in ecology.Science, 361(6406):eaat6412, 2018
2018
-
[14]
Morozov, K
A. Morozov, K. Abbott, K. Cuddington, T. Francis, G. Gellner, A. Hastings, and M. L. Zeeman. Long transients in ecology: Theory and applications.Physics of Life Reviews, 32:1–40, 2020
2020
-
[15]
Morozov, U
A. Morozov, U. Feudel, A. Hastings, K. C. Abbott, K. Cuddington, C. M. Heggerud, and S. Petro- vskii. Long-living transients in ecological models: Recent progress, new challenges, and open questions.Physics of Life Reviews, 51:423–441, 2024
2024
-
[16]
D. Koch, A. Nandan, G. Ramesan, I. Tyukin, A. Gorban, and A. Koseska. Ghost channels and ghost cycles guiding long transients in dynamical systems.Physical Review Letters, 133(4):047202, 2024
2024
-
[17]
C. S. Holling. Resilience and stability of ecological systems.Annual Review of Ecology and Systematics, 4:1–23, 1973
1973
-
[18]
C. S. Holling. Engineering resilience versus ecological resilience. In Peter Schultze, editor,Engi- neering within Ecological Constraints, pages 31–43. 1996. 25
1996
-
[19]
L. H. Gunderson, C. R. Allen, and C. S. Holling, editors.Foundations of Ecological Resilience. Island Press, 2012
2012
-
[20]
Krakovsk´ a, C
H. Krakovsk´ a, C. Kuehn, and I. P. Longo. Resilience of dynamical systems.European Journal of Applied Mathematics, 35(1):155–200, 2024
2024
-
[21]
K. Meyer. A mathematical review of resilience in ecology.Natural Resource Modeling, 29(3):339– 352, 2016
2016
-
[22]
J. M. Reed, B. E. Wolfe, and L. M. Romero. Is resilience a unifying concept for the biological sciences?iScience, 2024
2024
-
[23]
C. Hui. Carrying capacity, population equilibrium, and environment’s maximal load.Ecological Modelling, 192:317–320, 2006. 26
2006
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.