Evolution of cooperation in the multiplex
Pith reviewed 2026-06-28 18:54 UTC · model grok-4.3
The pith
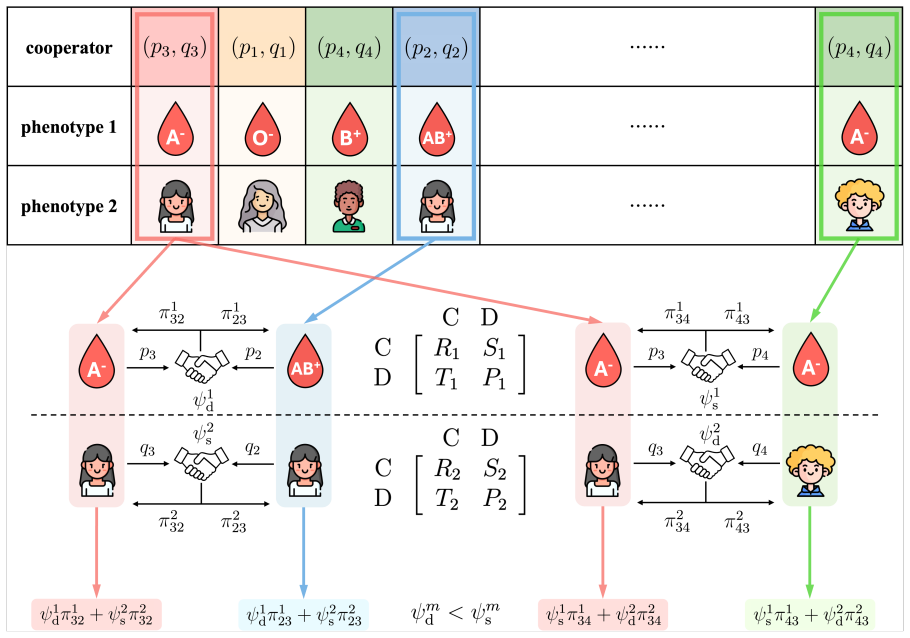
In multiplex networks with multi-phenotype homophily, selection for cooperation reduces to independent layer-specific σ-rules based on local payoffs, phenotype counts, and mutation rates.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
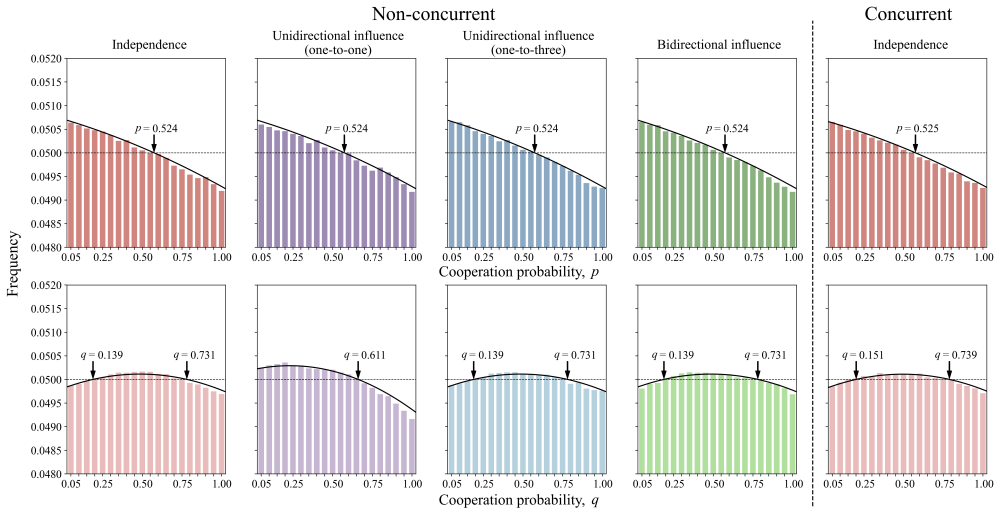
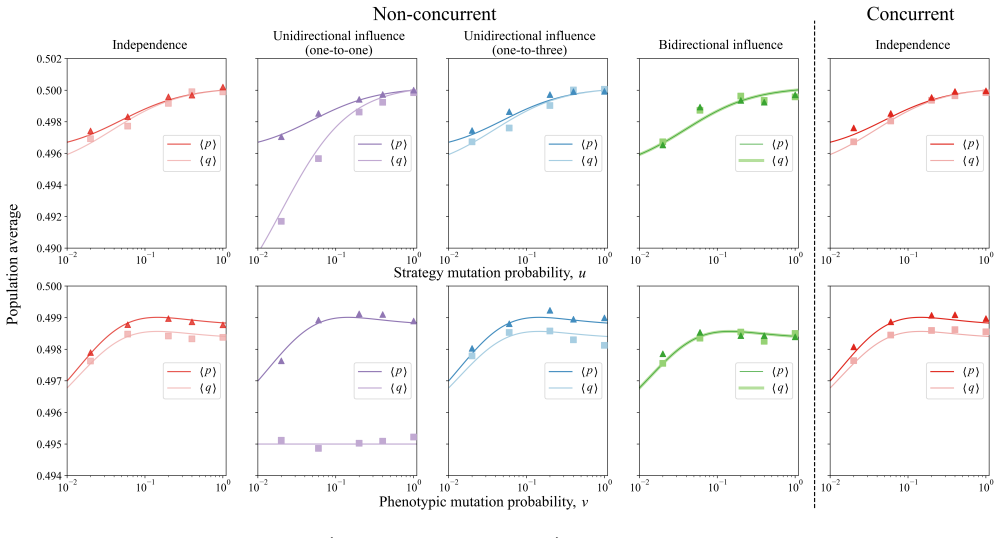
Despite fitness contributions being integrated across layers, the conditions for natural selection to favor cooperation resolve into layer-specific σ-rules that depend only on the local payoff structure, the effective number of phenotypes, and the mutation rates; phenotypic diversity fosters cooperation by partitioning populations into assortative niches, and in finite populations intensifying the prisoner's dilemma shifts the dependence of cooperation on strategy mutation from monotonically decreasing, through U-shaped, to monotonically increasing.
What carries the argument
layer-specific σ-rules under multi-phenotype homophily, which separate the selection condition for each phenotypic trait even when fitness is summed across layers.
If this is right
- Cooperation is favored precisely when each layer independently satisfies its own σ-rule.
- Increasing the number of phenotypic traits partitions the population into more assortative niches and thereby raises the chance that cooperation evolves.
- The shape of the relationship between cooperation level and strategy mutation rate is controlled by the intensity of the dilemma in finite populations.
Where Pith is reading between the lines
- The same separation into layer-specific rules may apply to any additive multi-context interaction, such as simultaneous economic and social payoffs.
- One could test whether observed cooperation rates in real multiplex data (e.g., online and offline contacts) match the predicted layer-by-layer σ conditions.
- The framework suggests that deliberately increasing phenotypic diversity in a population could be used as a control parameter to promote cooperation without changing payoffs.
Load-bearing premise
Fitness contributions from different layers combine in a manner that still permits the emergence of purely layer-specific selection conditions without additional cross-layer terms dominating.
What would settle it
A direct computation or simulation in a two-layer multiplex showing that the overall fixation probability of cooperators deviates from the product or combination of the individual layer σ-rules when mutation rates and payoff matrices are held fixed.
Figures

read the original abstract
Across biological and social systems, cooperation often depends on phenotypic cues rather than random encounters. To account for real-world interactions unfolding across multiple, simultaneous dimensions, here we develop a general framework for the evolution of cooperation in multiplex networks governed by multi-phenotype homophily. We derive analytical conditions for natural selection to favor cooperation across phenotypic traits that are independent or exhibit epistasis and under different modes of mutation coupling. Despite the integration of fitness across layers, the conditions for cooperation resolve into layer-specific $\sigma$-rules, depending only on the local payoff structure, the effective number of phenotypes, and the mutation rates. We show that phenotypic diversity fosters cooperation by partitioning populations into assortative niches. Furthermore, in finite populations, intensifying the prisoner's dilemma shifts the dependence of cooperation on strategy mutation from monotonically decreasing, through U-shaped, to monotonically increasing. Our work provides a unified account of how multi-phenotype homophily underpins the evolutionary dynamics of cooperation in heterogeneous populations.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a general framework for the evolution of cooperation in multiplex networks governed by multi-phenotype homophily. It derives analytical conditions for natural selection to favor cooperation across phenotypic traits that are independent or exhibit epistasis and under different modes of mutation coupling. Despite fitness integration across layers, the conditions resolve into layer-specific σ-rules depending only on the local payoff structure, the effective number of phenotypes, and the mutation rates. Phenotypic diversity is shown to foster cooperation by partitioning populations into assortative niches. In finite populations, intensifying the prisoner's dilemma shifts the dependence of cooperation on strategy mutation from monotonically decreasing, through U-shaped, to monotonically increasing.
Significance. If the derivations hold, the work provides a unified analytical account of multi-phenotype homophily in the evolutionary dynamics of cooperation in heterogeneous populations. The reduction to layer-specific σ-rules despite cross-layer fitness integration, together with the explicit results on mutation-rate dependence and the role of phenotypic diversity, constitute clear strengths. The analytical character of the conditions and the specific, falsifiable predictions on how prisoner's dilemma intensity alters mutation dependence are particularly valuable.
minor comments (3)
- The introduction would benefit from a brief explicit statement of the σ-rule from prior single-layer work (with citation) before introducing the multiplex extension, to aid readers new to the formalism.
- Notation for the effective number of phenotypes and the mutation-coupling modes should be defined once in a dedicated subsection or table rather than introduced piecemeal across sections.
- Figure captions for the finite-population simulations should state the exact parameter values used for mutation rates and population size so that the U-shaped regime can be reproduced directly from the text.
Simulated Author's Rebuttal
We thank the referee for their positive summary of our manuscript, accurate capture of the key results on layer-specific σ-rules under multi-phenotype homophily, and recommendation for minor revision. No specific major comments were raised in the report.
Circularity Check
No significant circularity
full rationale
The paper derives analytical conditions showing that multi-layer fitness integration resolves into layer-specific σ-rules depending only on local payoff structure, effective phenotype number, and mutation rates. No load-bearing steps reduce by construction to inputs: the separation is presented as a derived mathematical outcome under the stated assumptions on independent/epistatic traits and mutation modes, with no self-definitional equations, fitted parameters renamed as predictions, or self-citation chains visible in the abstract or claim descriptions. The framework is self-contained against external benchmarks in evolutionary game theory on networks.
Axiom & Free-Parameter Ledger
free parameters (2)
- mutation rates
- effective number of phenotypes
axioms (1)
- domain assumption Standard evolutionary game theory assumptions including weak selection and birth-death updating on networks.
Reference graph
Works this paper leans on
-
[1]
The evolution of homophily.Scientific reports, 2(1):845, 2012
Feng Fu, Martin A Nowak, Nicholas A Christakis, and James H Fowler. The evolution of homophily.Scientific reports, 2(1):845, 2012
2012
-
[2]
Random processes in genetics
Patrick Alfred Pierce Moran. Random processes in genetics. InMathematical proceedings of the cambridge philosophical society, volume 54, pages 60–71. Cambridge University Press, 1958
1958
-
[3]
Patrick Alfred Pierce Moran.The statistical processes of evolutionary theory.Clarendon Press, 1962
1962
-
[4]
Evolution and the theory of games
John Maynard Smith. Evolution and the theory of games. InDid Darwin get it right? Essays on games, sex and evolution, pages 202–215. Springer, 1982
1982
-
[5]
Cambridge university press, 1998
Josef Hofbauer and Karl Sigmund.Evolutionary games and population dynamics. Cambridge university press, 1998
1998
-
[6]
Evolution of cooperation by phenotypic similarity.Proceedings of the National Academy of Sciences, 106(21):8597–8600, 2009
Tibor Antal, Hisashi Ohtsuki, John Wakeley, Peter D Taylor, and Martin A Nowak. Evolution of cooperation by phenotypic similarity.Proceedings of the National Academy of Sciences, 106(21):8597–8600, 2009
2009
-
[7]
Mutation-selection equilibrium in games with multiple strategies.Journal of theoretical biol- ogy, 258(4):614–622, 2009
Tibor Antal, Arne Traulsen, Hisashi Ohtsuki, Corina E Tarnita, and Martin A Nowak. Mutation-selection equilibrium in games with multiple strategies.Journal of theoretical biol- ogy, 258(4):614–622, 2009
2009
-
[8]
Multiple strategies in structured populations.Proceedings of the National Academy of Sciences, 108(6):2334–2337, 2011
Corina E Tarnita, Nicholas Wage, and Martin A Nowak. Multiple strategies in structured populations.Proceedings of the National Academy of Sciences, 108(6):2334–2337, 2011
2011
-
[9]
Roberts & Company Publishers, 2008
John H Wakeley.Coalescent theory: an introduction. Roberts & Company Publishers, 2008. 32
2008
-
[10]
Strategy selection in structured populations.Journal of theoretical biology, 259(3):570–581, 2009
Corina E Tarnita, Hisashi Ohtsuki, Tibor Antal, Feng Fu, and Martin A Nowak. Strategy selection in structured populations.Journal of theoretical biology, 259(3):570–581, 2009
2009
-
[11]
Genome-wide association study identifies variants in the abo locus associated with susceptibility to pancreatic cancer.Nature genetics, 41(9):986–990, 2009
Laufey Amundadottir, Peter Kraft, Rachael Z Stolzenberg-Solomon, Charles S Fuchs, Glo- ria M Petersen, Alan A Arslan, H Bas Bueno-de Mesquita, Myron Gross, Kathy Helzlsouer, Eric J Jacobs, et al. Genome-wide association study identifies variants in the abo locus associated with susceptibility to pancreatic cancer.Nature genetics, 41(9):986–990, 2009
2009
-
[12]
The rh blood group system: a review.Blood, The Journal of the American Society of Hematology, 95(2):375–387, 2000
Neil D Avent and Marion E Reid. The rh blood group system: a review.Blood, The Journal of the American Society of Hematology, 95(2):375–387, 2000
2000
-
[13]
Model-based prediction of human hair color using dna variants.Human genetics, 129(4):443–454, 2011
Wojciech Branicki, Fan Liu, Kate van Duijn, Jolanta Draus-Barini, Ewelina Po´ spiech, Susan Walsh, Tomasz Kupiec, Anna Wojas-Pelc, and Manfred Kayser. Model-based prediction of human hair color using dna variants.Human genetics, 129(4):443–454, 2011
2011
-
[14]
Genetic determinants of hair, eye and skin pigmentation in europeans.Nature genetics, 39(12):1443–1452, 2007
Patrick Sulem, Daniel F Gudbjartsson, Simon N Stacey, Agnar Helgason, Thorunn Rafnar, Kristinn P Magnusson, Andrei Manolescu, Ari Karason, Arnar Palsson, Gudmar Thorleifsson, et al. Genetic determinants of hair, eye and skin pigmentation in europeans.Nature genetics, 39(12):1443–1452, 2007
2007
-
[15]
Deleterious mutations, variable epistatic inter- actions, and the evolution of recombination.Theoretical population biology, 51(2):134–147, 1997
Sarah Perin Otto and Marcus W Feldman. Deleterious mutations, variable epistatic inter- actions, and the evolution of recombination.Theoretical population biology, 51(2):134–147, 1997
1997
-
[16]
Integrated phenotypes: understanding trait covariation in plants and animals.Philosophical Transactions of the Royal Society B: Biological Sciences, 369(1649):20130245, 2014
W Scott Armbruster, Christophe P´ elabon, Geir H Bolstad, and Thomas F Hansen. Integrated phenotypes: understanding trait covariation in plants and animals.Philosophical Transactions of the Royal Society B: Biological Sciences, 369(1649):20130245, 2014
2014
-
[17]
Genetic studies of body mass index yield new insights for obesity biology.Nature, 518(7538):197–206, 2015
Adam E Locke, Bratati Kahali, Sonja I Berndt, Anne E Justice, Tune H Pers, Felix R Day, Corey Powell, Sailaja Vedantam, Martin L Buchkovich, Jian Yang, et al. Genetic studies of body mass index yield new insights for obesity biology.Nature, 518(7538):197–206, 2015
2015
-
[18]
Leaf, stem and root tissue strategies across 758 neotropical tree species.Functional ecology, 26(5):1153–1161, 2012
Claire Fortunel, Paul VA Fine, and Christopher Baraloto. Leaf, stem and root tissue strategies across 758 neotropical tree species.Functional ecology, 26(5):1153–1161, 2012
2012
-
[19]
The concept and definition of dominance in animal behaviour.Behaviour, 125(3-4):283–313, 1993
Carlos Drews. The concept and definition of dominance in animal behaviour.Behaviour, 125(3-4):283–313, 1993
1993
-
[20]
The evolution of a social mind
Dorothy L Cheney and Robert M Seyfarth. The evolution of a social mind. InBaboon metaphysics. University of Chicago Press, 2007
2007
-
[21]
Noah D Simons, Vasiliki Michopoulos, Mark Wilson, Luis B Barreiro, and Jenny Tung. Ago- nism and grooming behaviour explain social status effects on physiology and gene regulation in rhesus macaques.Philosophical Transactions of the Royal Society B: Biological Sciences, 377(1845):20210132, 2022
2022
-
[22]
David Mech and Luigi Boitani
L. David Mech and Luigi Boitani. Wolf social ecology. InWolves: Behavior, ecology and conservation. University of Chicago Press, 2003
2003
-
[23]
Heritabilities and genetic correlations of body weights and feather length in growing muscovy selected in taiwan.British Poultry Science, 40(5):605–612, 1999
YH Hu, Jean Paul Poivey, Roger Rouvier, CT Wang, and Chein Tai. Heritabilities and genetic correlations of body weights and feather length in growing muscovy selected in taiwan.British Poultry Science, 40(5):605–612, 1999. 33
1999
-
[24]
The end of materialism? InBritish Social Attitudes: The Fifteenth Report, pages 125–148
Caroline Bryson and John Curtice. The end of materialism? InBritish Social Attitudes: The Fifteenth Report, pages 125–148. Ashgate, Aldershot, 1998
1998
-
[25]
Coevolutionary dynamics of phenotypic diversity and contingent cooperation.PLoS computational biology, 13(1):e1005363, 2017
Te Wu, Long Wang, and Feng Fu. Coevolutionary dynamics of phenotypic diversity and contingent cooperation.PLoS computational biology, 13(1):e1005363, 2017
2017
-
[26]
The role of diversity in the evolution of cooperation.Journal of theoretical biology, 299:88–96, 2012
Francisco C Santos, Flavio L Pinheiro, Tom Lenaerts, and Jorge M Pacheco. The role of diversity in the evolution of cooperation.Journal of theoretical biology, 299:88–96, 2012
2012
-
[27]
Fisher.The Genetical Theory of Natural Selection
Ronald A. Fisher.The Genetical Theory of Natural Selection. Clarendon Press, Oxford, 1930
1930
-
[28]
Evolution in mendelian populations.Genetics, 16(2):97, 1931
Sewall Wright. Evolution in mendelian populations.Genetics, 16(2):97, 1931
1931
-
[29]
Evolutionary dynamics in struc- tured populations.Philosophical Transactions of the Royal Society B: Biological Sciences, 365(1537):19–30, 2010
Martin A Nowak, Corina E Tarnita, and Tibor Antal. Evolutionary dynamics in struc- tured populations.Philosophical Transactions of the Royal Society B: Biological Sciences, 365(1537):19–30, 2010
2010
-
[30]
Evolutionary dynamics in set structured populations.Proceedings of the National Academy of Sciences, 106(21):8601– 8604, 2009
Corina E Tarnita, Tibor Antal, Hisashi Ohtsuki, and Martin A Nowak. Evolutionary dynamics in set structured populations.Proceedings of the National Academy of Sciences, 106(21):8601– 8604, 2009
2009
-
[31]
Evolution of cooperation by phenotypic similarity.Proceedings of the National Academy of Sciences, 106(21):8597–8600, 2009
Tibor Antal, Hisashi Ohtsuki, John Wakeley, Peter D Taylor, and Martin A Nowak. Evolution of cooperation by phenotypic similarity.Proceedings of the National Academy of Sciences, 106(21):8597–8600, 2009. 34
2009
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.