Inferring long-range interactions between immune and tumor cells -- pitfalls and (partial) solutions
Pith reviewed 2026-05-24 16:48 UTC · model grok-4.3
The pith
A turning angle probability model correctly infers cell-cell interactions even when migration behavior switches over time.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The distance-dependent turning angle probability model recovers the correct interaction parameters even with temporally switching cell migration, while the force model detects spurious interactions in independent cells that change migration behavior.
What carries the argument
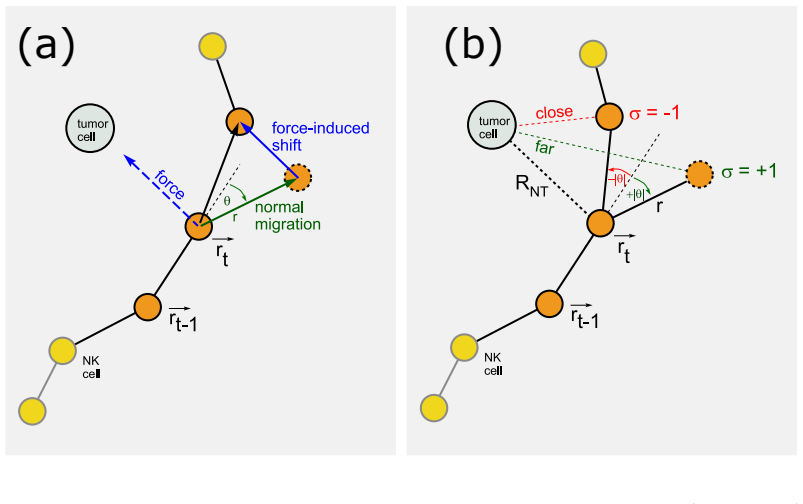
Distance-dependent turning angle probability model, which fits the probabilities of positive and negative turning angles of an immune cell as a function of its distance to the nearest tumor cell via maximum likelihood.
Load-bearing premise
Simulated trajectories with constant or temporally switching migration properties adequately represent the confounding factors and data characteristics present in actual experimental cell recordings.
What would settle it
Generate trajectories of non-interacting cells programmed to switch migration directions or speeds at random times, then verify whether the turning angle model returns near-zero interaction strength while the force model returns positive strength.
Figures






read the original abstract
Upcoming immunotherapies for cancer treatment rely on the ability of the immune system to detect and eliminate tumors in the body. A highly simplified version of this process can be studied in a Petri dish: starting with a random distribution of immune and tumor cells, it can be observed in detail how individual immune cells migrate towards nearby tumor cells, establish contact, and attack. Nevertheless, it remains unclear whether the immune cells find their targets by chance, or if they approach them 'on purpose', using remote sensing mechanisms such as chemotaxis. In this work, we present methods to infer the strength and range of long-range cell-cell interactions from time-lapse recorded cell trajectories, using a maximum likelihood method to fit the model parameters. First, we model the interactions as a distance-dependent 'force' that attracts immune cells towards their nearest tumor cell. While this approach correctly recovers the interaction parameters of simulated cells with constant migration properties, it detects spurious interactions in the case of independent cells that spontaneously change their migration behavior over time. We therefore use an alternative approach that models the interactions by distance-dependent probabilities for positive and negative turning angles of the migrating immune cell. We demonstrate that the latter approach finds the correct interaction parameters even with temporally switching cell migration.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that a distance-dependent force model for inferring long-range immune-tumor cell interactions from trajectories recovers true parameters in simulations with constant migration properties but produces spurious interactions when cells independently switch migration behavior over time; an alternative model using distance-dependent probabilities for positive/negative turning angles recovers the correct parameters even under temporal switching, as demonstrated via maximum-likelihood fitting on forward-simulated trajectories with known ground truth.
Significance. If the turning-angle approach holds under more realistic conditions, the work identifies a substantive pitfall in force-based inference methods commonly used for cell trajectory analysis and provides a concrete alternative. Credit is due for the use of simulations with independently set interaction parameters and behavioral switches, which supplies an external (non-circular) check on recovery rather than reducing to fitted quantities by construction.
major comments (1)
- [Simulation validation (abstract and results)] The central claim—that the turning-angle probability model recovers true interaction parameters under temporally switching migration while the force model yields false positives—rests on the assumption that the simulated trajectories adequately capture the confounding factors in real experimental cell recordings. No quantitative match is shown between simulated and experimental turning-angle distributions, speed histograms, or persistence statistics (see abstract and simulation results).
Simulated Author's Rebuttal
We thank the referee for their constructive review and recommendation. Our response to the single major comment is provided below. The manuscript uses controlled simulations with known ground truth to demonstrate a methodological pitfall and a proposed solution; we defend that this does not require quantitative matching to experimental distributions.
read point-by-point responses
-
Referee: [Simulation validation (abstract and results)] The central claim—that the turning-angle probability model recovers true interaction parameters under temporally switching migration while the force model yields false positives—rests on the assumption that the simulated trajectories adequately capture the confounding factors in real experimental cell recordings. No quantitative match is shown between simulated and experimental turning-angle distributions, speed histograms, or persistence statistics (see abstract and simulation results).
Authors: We respectfully disagree that the central claim requires quantitative matching between simulated and experimental statistics. The manuscript's core contribution is a controlled demonstration, via forward simulations with independently specified ground-truth interaction parameters and behavioral switches, that the force-based model produces spurious detections while the turning-angle probability model recovers the true parameters. This isolates temporal heterogeneity as a confounding factor without circularity. The simulations incorporate persistence, speed variation, and distance-dependent turning to capture the relevant mechanism, but the validation is external (recovery of known parameters) rather than a claim to reproduce any specific experimental dataset. The abstract and results sections make no assertion that the simulated trajectories match real recordings; the focus is methodological robustness. Adding such matches would require new experimental data and is outside the current scope, though we would be willing to expand the discussion of simulation parameterization if helpful. revision: no
Circularity Check
No significant circularity; validation uses independent forward simulations
full rationale
The paper's derivation chain consists of defining two alternative interaction models (force-based and turning-angle probability) and testing parameter recovery on simulated trajectories. Interaction parameters are set independently in the simulations before fitting, and recovery success is measured against those known values. This constitutes an external check rather than any reduction of outputs to inputs by construction. No self-definitional equations, fitted-input predictions, or load-bearing self-citations appear in the provided text. The central claim therefore remains independent of the fitting procedure itself.
Axiom & Free-Parameter Ledger
free parameters (2)
- interaction strength
- interaction range
axioms (2)
- standard math Maximum likelihood estimation recovers true parameters when data are generated from the assumed interaction model.
- domain assumption Cell migration can be decomposed into either distance-dependent forces or distance-dependent turning probabilities plus independent behavioral switches.
Reference graph
Works this paper leans on
-
[1]
Immunobiology: the immune system in health and disease , volume 7
Charles A Janeway, Paul Travers, Mark Walport, Mark Shlomchik, et al. Immunobiology: the immune system in health and disease , volume 7. Current Biology London, 1996. 12
work page 1996
-
[2]
Pathogen recognition by the innate immune system
Himanshu Kumar, Taro Kawai, and Shizuo Akira. Pathogen recognition by the innate immune system. International reviews of immunology , 30(1):16–34, 2011
work page 2011
-
[3]
Manfred Schuster, Andreas Nechansky, and Ralf Kircheis. Cancer immunotherapy. Biotech- nology Journal: Healthcare Nutrition Technology , 1(2):138–147, 2006
work page 2006
-
[4]
Decade in reviewcancer immunotherapy: entering the mainstream of cancer treatment
Steven A Rosenberg. Decade in reviewcancer immunotherapy: entering the mainstream of cancer treatment. Nature Reviews Clinical Oncology, 11(11):630, 2014
work page 2014
-
[5]
Breathing new life into immunother- apy: review of melanoma, lung and kidney cancer
Charles G Drake, Evan J Lipson, and Julie R Brahmer. Breathing new life into immunother- apy: review of melanoma, lung and kidney cancer. Nature reviews Clinical oncology, 11(1):24, 2014
work page 2014
-
[6]
Distinct migration and contact dynamics of resting and il-2-activated human natural killer cells
Per Erik Olofsson, Elin Forslund, Bruno Vanherberghen, Ksenia Chechet, Oscar Mickelin, Alexander Rivera Ahlin, Tobias Everhorn, and Bj¨ orn¨Onfelt. Distinct migration and contact dynamics of resting and il-2-activated human natural killer cells. Frontiers in immunology , 5:80, 2014
work page 2014
-
[7]
Bystander cells enhance nk cytotoxic efficiency by reducing search time
Xiao Zhou, Renping Zhao, Karsten Schwarz, Matthieu Mangeat, Eva C Schwarz, Mohamed Hamed, Ivan Bogeski, Volkhard Helms, Heiko Rieger, and Bin Qu. Bystander cells enhance nk cytotoxic efficiency by reducing search time. Scientific reports, 7:44357, 2017
work page 2017
- [8]
-
[9]
Edward A Codling, Michael J Plank, and Simon Benhamou. Random walk models in biology. Journal of the Royal society interface , 5(25):813–834, 2008
work page 2008
-
[10]
Volkmar Heinrich, Wooten D Simpson III, and Emmet A Francis. Analytical prediction of the spatiotemporal distribution of chemoattractants around their source: Theory and application to complement-mediated chemotaxis. Frontiers in immunology, 8:578, 2017
work page 2017
-
[11]
Superstatistical analysis and modelling of heterogeneous random walks
Claus Metzner, Christoph Mark, Julian Steinwachs, Lena Lautscham, Franz Stadler, and Ben Fabry. Superstatistical analysis and modelling of heterogeneous random walks. Nature communications, 6(May):7516, jun 2015
work page 2015
-
[12]
Principles of efficient chemotactic pursuit
Claus Metzner. Principles of efficient chemotactic pursuit. arXiv:1902.10589, 2019
work page internal anchor Pith review Pith/arXiv arXiv 1902
-
[13]
Detecting long-range attraction between migrating cells based on p-value distributions
Claus Metzner. Detecting long-range attraction between migrating cells based on p-value distributions. arXiv:1906.08481, 2019. 13 ADDITIONAL INFORMA TION Author contributions statement: CM has devised the study, developed the simulation programs, performed and interpreted the simulations, and wrote the paper. Funding: This work was funded by the Grant ME1...
work page internal anchor Pith review Pith/arXiv arXiv 1906
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.