Dual Mechanisms for Heterogeneous Responses of Inspiratory Neurons to Noradrenergic Modulation
Pith reviewed 2026-05-19 02:46 UTC · model grok-4.3
The pith
Norepinephrine modulates inspiratory neurons through simultaneous changes in calcium-activated current conductance and IP3 to produce cell-specific bursting patterns.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
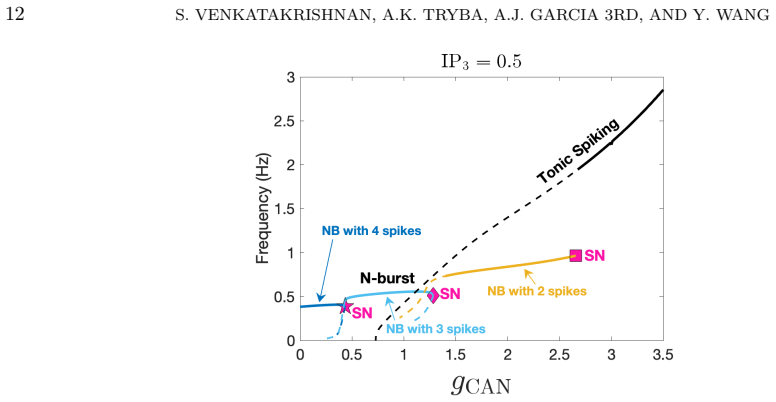
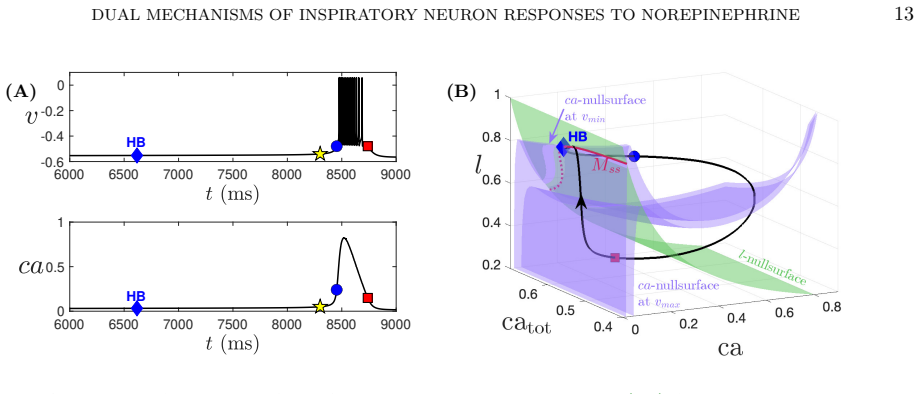
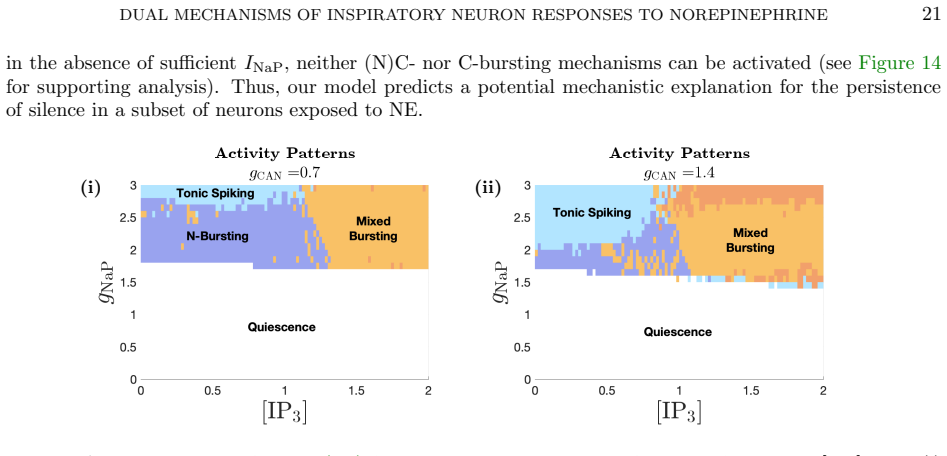
By modulating g_CAN and IP3 in a computational model of preBötC neurons, norepinephrine differentially affects NaP-dependent and CAN-dependent bursting neurons. The dual mechanism is critical for inducing conditional bursting, and specific parameter regimes keep silent neurons inactive even in the presence of NE. Dynamical systems analysis reveals how NE modulates burst frequency and duration differently depending on the underlying bursting mechanism, and these outcomes align with previously reported experimental findings.
What carries the argument
Dual modulation of the calcium-activated nonspecific cationic current conductance (g_CAN) and inositol-triphosphate (IP3) concentration, which together reproduce the experimentally observed heterogeneous responses to norepinephrine.
If this is right
- Conditional bursting occurs only when both g_CAN and IP3 are modulated together rather than either parameter alone.
- Silent neurons remain inactive across identifiable ranges of the two parameters even when norepinephrine is present.
- Norepinephrine produces opposite or distinct changes in burst frequency versus burst duration depending on whether the neuron uses NaP-dependent or CAN-dependent mechanisms.
- The modeled outcomes match the differential effects reported in experimental studies of norepinephrine on preBötC neurons.
Where Pith is reading between the lines
- The same dual-parameter approach could be tested on other neuromodulators to see whether similar mechanisms explain their cell-specific actions in the respiratory network.
- Mapping the parameter regimes that preserve silence might help predict conditions under which breathing rhythm stays unchanged despite neuromodulatory input.
- Incorporating these single-neuron mechanisms into network models could clarify how heterogeneous responses scale up to shape overall inspiratory rhythm stability and flexibility.
Load-bearing premise
The experimentally observed effects of norepinephrine can be captured by modulating only g_CAN and IP3 while holding all other cellular properties fixed or scaling them uniformly.
What would settle it
An experiment that shows norepinephrine must change additional cellular properties beyond g_CAN and IP3 to produce the full range of differential effects on preBötC neuron bursting would indicate that the dual-parameter model is incomplete.
Figures











read the original abstract
Respiration is an essential involuntary function necessary for survival. This poses a challenge for the control of breathing. The preB\"otzinger complex (preB\"otC) is a heterogeneous neuronal network responsible for driving the inspiratory rhythm. While neuromodulators such as norepinephrine (NE) allow it to be both robust and flexible for all living beings to interact with their environment, the basis for how neuromodulation impacts neuron-specific properties remains poorly understood. In this work, we examine how NE influences different preB\"otC neuronal subtypes by modeling its effects through modulating two key parameters: calcium-activated nonspecific cationic current gating conductance ($g_{\rm CAN}$) and inositol-triphosphate ($\rm IP_3$), guided by experimental studies. Our computational model captures the experimentally observed differential effects of NE on distinct preB\"otC bursting patterns. We show that this dual mechanism is critical for inducing conditional bursting and identify specific parameter regimes where silent neurons remain inactive in the presence of NE. Furthermore, using methods of dynamical systems theory, we uncover the mechanisms by which NE differentially modulates burst frequency and duration in NaP-dependent and CAN-dependent bursting neurons. These results align well with previously reported experimental findings and provide a deeper understanding of cell-specific neuromodulatory responses within the respiratory network.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper develops a computational model of preBötC inspiratory neurons to examine norepinephrine (NE) effects on heterogeneous bursting. Guided by experiments, it modulates only g_CAN and IP3 to reproduce differential NE responses across NaP-dependent and CAN-dependent subtypes, claims this dual mechanism is critical for conditional bursting, identifies parameter regimes where silent neurons remain inactive, and applies dynamical systems analysis to explain NE-induced changes in burst frequency and duration.
Significance. If the modeling assumptions hold, the work offers mechanistic insight into cell-specific neuromodulation within the respiratory rhythm generator and demonstrates how dynamical systems methods can link parameter changes to specific dynamical features such as burst metrics. This could inform broader questions of network robustness and flexibility under neuromodulatory control.
major comments (2)
- [Abstract] Abstract: The assertion that the dual mechanism (modulation of g_CAN and IP3) is 'critical' for inducing conditional bursting and for the observed differential effects is not supported by any comparison to single-parameter modulation, alternative NE targets (e.g., leak or NaP conductances), or sensitivity analysis. Without these, the necessity of the two-parameter restriction remains untested and the claim risks being an artifact of the restricted parameter space.
- [Abstract] Abstract / parameter selection: Ranges for g_CAN and IP3 are stated to be 'guided by experimental studies,' yet the manuscript provides no explicit check that these ranges were chosen independently of the model outputs or that the heterogeneity and conditional bursting emerge robustly outside the fitted regimes. This raises a potential circularity concern for the central claim that the dual mechanism explains the experimental observations.
minor comments (1)
- [Abstract] Abstract: The statement that results 'align well with previously reported experimental findings' would benefit from citing the specific studies and providing at least qualitative or quantitative matches for burst frequency, duration, or conditional bursting incidence.
Simulated Author's Rebuttal
We thank the referee for their constructive comments. We address each major comment below and indicate where revisions will be made to the manuscript.
read point-by-point responses
-
Referee: [Abstract] Abstract: The assertion that the dual mechanism (modulation of g_CAN and IP3) is 'critical' for inducing conditional bursting and for the observed differential effects is not supported by any comparison to single-parameter modulation, alternative NE targets (e.g., leak or NaP conductances), or sensitivity analysis. Without these, the necessity of the two-parameter restriction remains untested and the claim risks being an artifact of the restricted parameter space.
Authors: We acknowledge that the word 'critical' implies a necessity that is not directly tested by comparisons to single-parameter changes or other potential NE targets. The dual modulation was selected because experimental literature specifically implicates NE effects on g_CAN and IP3 in preBötC neurons. To address this point, we will add supplementary simulations comparing single versus dual modulation and include a brief sensitivity analysis. We will also revise the abstract language to 'this dual mechanism enables' rather than 'is critical for' to more accurately reflect the presented results. revision: yes
-
Referee: [Abstract] Abstract / parameter selection: Ranges for g_CAN and IP3 are stated to be 'guided by experimental studies,' yet the manuscript provides no explicit check that these ranges were chosen independently of the model outputs or that the heterogeneity and conditional bursting emerge robustly outside the fitted regimes. This raises a potential circularity concern for the central claim that the dual mechanism explains the experimental observations.
Authors: The ranges were taken from published experimental measurements of NE-induced changes in CAN current and IP3 levels in respiratory neurons and were not tuned to match specific model outputs. To make this independence explicit and address the robustness concern, we will add a dedicated paragraph in the Methods section citing the experimental sources and include additional figures or text showing that the key qualitative behaviors (heterogeneity and conditional bursting) persist when parameters are varied modestly outside the primary ranges. revision: yes
Circularity Check
No significant circularity; derivation chain is self-contained
full rationale
The paper constructs a computational model of preBötC neurons, selects modulations to g_CAN and IP3 guided by prior experimental literature, then applies dynamical systems analysis to derive mechanisms for conditional bursting and differential effects on frequency/duration. No step reduces by construction to the inputs: the criticality claim follows from explicit simulation of the two-parameter space and phase-plane analysis rather than tautological renaming or refitting of observed outputs. No self-citation load-bearing uniqueness theorems, ansatz smuggling, or fitted-input predictions are present. The work remains independent of its own parameter choices because the dynamical mechanisms are falsifiable against the model's equations and align with but are not forced by the experimental guidance.
Axiom & Free-Parameter Ledger
free parameters (2)
- g_CAN modulation factor
- IP3 modulation factor
axioms (1)
- domain assumption The preBötC neuron model equations and baseline parameters from prior literature remain valid when only g_CAN and IP3 are varied.
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
modeling its effects through modulating two key parameters: calcium-activated nonspecific cationic current gating conductance (g_CAN) and inositol-triphosphate (IP3)
-
IndisputableMonolith/Foundation/AlexanderDuality.leanalexander_duality_circle_linking unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
using methods of dynamical systems theory, we uncover the mechanisms by which NE differentially modulates burst frequency and duration
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
N. A. Baertsch, N. E. Bush, N. J. Burgraff, and J.-M. Ramirez , Dual mechanisms of opioid-induced respiratory depression in the inspiratory rhythm-generating network , Elife, 10 (2021), p. e67523
work page 2021
-
[2]
N. A. Baertsch and J.-M. Ramirez , Insights into the dynamic control of breathing revealed through cell-type-specific responses to substance p , Elife, 8 (2019), p. e51350
work page 2019
-
[3]
H. Baldemir, D. A vitabile, and K. Tsaneva-Atanasova , Pseudo-plateau bursting and mixed-mode oscillations in a model of developing inner hair cells , Communications in Nonlinear Science and Numerical Simulation, 80 (2020), p. 104979
work page 2020
-
[4]
F. Ben-Mabrouk and A. K. Tryba , Substance p modulation of trpc3/7 channels improves respiratory rhythm regularity and ican-dependent pacemaker activity, European Journal of Neuroscience, 31 (2010), pp. 1219–1232
work page 2010
-
[5]
R. Bertram, M. J. Butte, T. Kiemel, and A. Sherman , Topological and phenomenological classification of bursting oscillations, Bulletin of mathematical biology, 57 (1995), pp. 413–439
work page 1995
-
[6]
R. Bertram, J. Rhoads, and W. P. Cimbora , A phantom bursting mechanism for episodic bursting , Bulletin of Math- ematical Biology, 70 (2008), pp. 1979–1993
work page 2008
-
[7]
B. M. Browe, Y.-J. Peng, J. Nanduri, N. R. Prabhakar, and A. J. Garcia III , Gasotransmitter modulation of hypoglossal motoneuron activity, Elife, 12 (2023), p. e81978
work page 2023
-
[8]
N. J. Burgraff, N. E. Bush, J. M. Ramirez, and N. A. Baertsch , Dynamic rhythmogenic network states drive differential opioid responses in the in vitro respiratory network , Journal of Neuroscience, 41 (2021), pp. 9919–9931
work page 2021
-
[9]
R. J. Butera, J. Rinzel, and J. C. Smith , Models of respiratory rhythm generation in the pre-botzinger complex. i. bursting pacemaker neurons, Journal of neurophysiology, 82 (1999), pp. 382–397
work page 1999
-
[10]
R. J. Butera, J. Rinzel, and J. C. Smith , Models of respiratory rhythm generation in the pre-botzinger complex. ii. populations of coupled pacemaker neurons, Journal of neurophysiology, 82 (1999), pp. 398–415
work page 1999
-
[11]
M. Chevalier, N. Toporikova, J. Simmers, and M. Thoby-Brisson , Development of pacemaker properties and rhyth- mogenic mechanisms in the mouse embryonic respiratory network , Elife, 5 (2016), p. e16125
work page 2016
-
[12]
G. M. Chou, N. E. Bush, R. S. Phillips, N. A. Baertsch, and K. D. Harris , Modeling effects of variable preb¨ otzinger complex network topology and cellular properties on opioid-induced respiratory depression and recovery , eneuro, 11 (2024)
work page 2024
-
[13]
J. M. Choy, F. A. Agahari, L. Li, and C. Stricker , Noradrenaline increases mepsc frequency in pyramidal cells in layer ii of rat barrel cortex via calcium release from presynaptic stores , Frontiers in cellular neuroscience, 12 (2018), p. 213
work page 2018
-
[14]
C. A. Del Negro, C. Morgado-V alle, J. A. Hayes, D. D. Mackay, R. W. Pace, E. A. Crowder, and J. L. Feldman, Sodium and calcium current-mediated pacemaker neurons and respiratory rhythm generation , Journal of Neuroscience, 25 (2005), pp. 446–453
work page 2005
-
[15]
M. Desroches, J. Guckenheimer, B. Krauskopf, C. Kuehn, H. M. Osinga, and M. Wechselberger , Mixed-mode oscillations with multiple time scales , SIAM Rev., 54 (2012), pp. 211–288
work page 2012
-
[16]
M. Desroches, T. J. Kaper, and M. Krupa , Mixed-mode bursting oscillations: Dynamics created by a slow passage through spike-adding canard explosion in a square-wave burster , Chaos, 23 (2013)
work page 2013
-
[17]
M. Desroches, J. Rinzel, and S. Rodrigues , Classification of bursting patterns: A tale of two ducks , PLoS Comput. Biol., 18 (2022)
work page 2022
-
[18]
R. R. Dhingra, P. M. MacF arlane, P. J. Thomas, J. F. Paton, and M. Dutschmann , Asymmetric neuromodulation in the respiratory network contributes to rhythm and pattern generation , bioRxiv, (2024), pp. 2024–11
work page 2024
-
[19]
A. Doi and J.-M. Ramirez , Neuromodulation and the orchestration of the respiratory rhythm , Respiratory physiology & neurobiology, 164 (2008), pp. 96–104. 32 S. VENKATAKRISHNAN, A.K. TRYBA, A.J. GARCIA 3RD, AND Y. WANG
work page 2008
-
[20]
J. R. Dunmyre, C. A. Del Negro, and J. E. Rubin , Interactions of persistent sodium and calcium-activated non- specific cationic currents yield dynamically distinct bursting regimes in a model of respiratory neurons , Journal of computational neuroscience, 31 (2011), pp. 305–328
work page 2011
-
[21]
J. Exton , Mechanisms involved in alpha-adrenergic phenomena , American Journal of Physiology-Endocrinology And Metabolism, 248 (1985), pp. E633–E647
work page 1985
-
[22]
N. Fenichel, Geometric singular perturbation theory for ordinary differential equations, Journal of differential equations, 31 (1979), pp. 53–98
work page 1979
-
[23]
G. D. Funk, J. C. Smith, and J. L. Feldman , Generation and transmission of respiratory oscillations in medullary slices: role of excitatory amino acids , Journal of neurophysiology, 70 (1993), pp. 1497–1515
work page 1993
-
[24]
A. J. Garcia III, S. Zanella, T. Dashevskiy, S. A. Khan, M. A. Khuu, N. R. Prabhakar, and J.-M. Ramirez , Chronic intermittent hypoxia alters local respiratory circuit function at the level of the preb¨ otzinger complex, Frontiers in neuroscience, 10 (2016), p. 4
work page 2016
-
[25]
M. Golubitsky, K. Josic, and T. J. Kaper , An unfolding theory approach to bursting in fast–slow systems , in Global analysis of dynamical systems, CRC Press, 2001, pp. 282–313
work page 2001
-
[26]
B. P. Guiard, M. El Mansari, Z. Merali, and P. Blier , Functional interactions between dopamine, serotonin and nor- epinephrine neurons: an in-vivo electrophysiological study in rats with monoaminergic lesions , International Journal of Neuropsychopharmacology, 11 (2008), pp. 625–639
work page 2008
-
[27]
R. Helliwell and W. Large , Alpha 1-adrenoceptor activation of a non-selective cation current in rabbit portal vein by 1, 2-diacyl-sn-glycerol., The Journal of Physiology, 499 (1997), pp. 417–428
work page 1997
-
[28]
A. J. Hill, J. M. Hinton, H. Cheng, Z. Gao, D. O. Bates, J. C. Hancox, P. D. Langton, and A. F. James , A trpc-like non-selective cation current activated by α1-adrenoceptors in rat mesenteric artery smooth muscle cells, Cell calcium, 40 (2006), pp. 29–40
work page 2006
-
[29]
A. L. Hodgkin and A. F. Huxley , A quantitative description of membrane current and its application to conduction and excitation in nerve , The Journal of physiology, 117 (1952), p. 500
work page 1952
-
[30]
E. M. Izhikevich , Neural excitability, spiking and bursting , International journal of bifurcation and chaos, 10 (2000), pp. 1171–1266
work page 2000
-
[31]
P. E. Jasinski, Y. I. Molkov, N. A. Shevtsova, J. C. Smith, and I. A. Rybak , Sodium and calcium mechanisms of rhythmic bursting in excitatory neural networks of the pre-b ¨ otzinger complex: a computational modelling study , European Journal of Neuroscience, 37 (2013), pp. 212–230
work page 2013
-
[32]
S. M. Johnson, J. C. Smith, G. D. Funk, and J. L. Feldman , Pacemaker behavior of respiratory neurons in medullary slices from neonatal rat, Journal of neurophysiology, 72 (1994), pp. 2598–2608
work page 1994
-
[33]
H. Koizumi and J. C. Smith , Persistent na+ and k+-dominated leak currents contribute to respiratory rhythm generation in the pre-b¨ otzinger complex in vitro, Journal of Neuroscience, 28 (2008), pp. 1773–1785
work page 2008
-
[34]
E. K ¨oksal Ers ¨oz, M. Desroches, A. Guillamon, J. Rinzel, and J. Tabak , Canard-induced complex oscillations in an excitatory network , Journal of mathematical biology, 80 (2020), pp. 2075–2107
work page 2020
- [35]
-
[36]
P. Nan, Y. W ang, V. Kirk, and J. E. Rubin , Understanding and distinguishing three-time-scale oscillations: Case study in a coupled morris–lecar system , SIAM journal on applied dynamical systems, 14 (2015), pp. 1518–1557
work page 2015
-
[37]
R. W. Pace, D. D. Mackay, J. L. Feldman, and C. A. Del Negro , Inspiratory bursts in the preb¨ otzinger complex depend on a calcium-activated non-specific cation current linked to glutamate receptors in neonatal mice, The Journal of physiology, 582 (2007), pp. 113–125
work page 2007
-
[38]
C. Park and J. E. Rubin , Cooperation of intrinsic bursting and calcium oscillations underlying activity patterns of model pre-b¨ otzinger complex neurons, Journal of Computational Neuroscience, 34 (2013), pp. 345–366
work page 2013
-
[39]
F. Pe ˜na and M.-A. Aguileta , Effects of riluzole and flufenamic acid on eupnea and gasping of neonatal mice in vivo , Neuroscience letters, 415 (2007), pp. 288–293
work page 2007
- [40]
-
[41]
N. A. Phan and Y. W ang , Mixed-mode oscillations in a three-timescale coupled morris–lecar system , Chaos: An Inter- disciplinary Journal of Nonlinear Science, 34 (2024)
work page 2024
-
[42]
R. S. Phillips, T. T. John, H. Koizumi, Y. I. Molkov, and J. C. Smith , Biophysical mechanisms in the mammalian respiratory oscillator re-examined with a new data-driven computational model , Elife, 8 (2019), p. e41555
work page 2019
-
[43]
R. S. Phillips and J. E. Rubin , Putting the theory into ‘burstlet theory’with a biophysical model of burstlets and bursts in the respiratory preb¨ otzinger complex, Elife, 11 (2022), p. e75713
work page 2022
-
[44]
J.-M. Ramirez, H. Koch, A. J. Garcia, A. Doi, and S. Zanella , The role of spiking and bursting pacemakers in the neuronal control of breathing, Journal of biological physics, 37 (2011), pp. 241–261
work page 2011
-
[45]
D. W. Richter, T. Manzke, B. Wilken, and E. Ponimaskin , Serotonin receptors: guardians of stable breathing, Trends in molecular medicine, 9 (2003), pp. 542–548
work page 2003
-
[46]
J. Rinzel , A formal classification of bursting mechanisms in excitable systems , in Mathematical Topics in Population Biology, Morphogenesis and Neurosciences: Proceedings of an International Symposium held in Kyoto, November 10–15, 1985, Springer, 1987, pp. 267–281
work page 1985
-
[47]
J. E. Rubin, J. A. Hayes, J. L. Mendenhall, and C. A. Del Negro , Calcium-activated nonspecific cation current and synaptic depression promote network-dependent burst oscillations , Proceedings of the National Academy of Sciences, 106 (2009), pp. 2939–2944. DUAL MECHANISMS OF INSPIRATORY NEURON RESPONSES TO NOREPINEPHRINE 33
work page 2009
-
[48]
X. Shao and J. Feldman , Acetylcholine modulates respiratory pattern: effects mediated by m3-like receptors in pre- botzinger complex inspiratory neurons, Journal of neurophysiology, 83 (2000), pp. 1243–1252
work page 2000
-
[49]
J. C. Smith, H. H. Ellenberger, K. Ballanyi, D. W. Richter, and J. L. Feldman , Pre-b¨ otzinger complex: a brainstem region that may generate respiratory rhythm in mammals , Science, 254 (1991), pp. 726–729
work page 1991
-
[50]
M. Thoby-Brisson and J.-M. Ramirez , Identification of two types of inspiratory pacemaker neurons in the isolated respiratory neural network of mice , Journal of neurophysiology, 86 (2001), pp. 104–112
work page 2001
-
[51]
N. Toporikova and R. J. Butera , Two types of independent bursting mechanisms in inspiratory neurons: an integrative model, Journal of computational neuroscience, 30 (2011), pp. 515–528
work page 2011
-
[52]
N. Toporikova and R. J. Butera Jr , Dynamics of neuromodulatory feedback determines frequency modulation in a reduced respiratory network: A computational study, Respiratory physiology & neurobiology, 185 (2013), pp. 582–592
work page 2013
-
[53]
N. Toporikova, M. Chevalier, and M. Thoby-Brisson , Sigh and eupnea rhythmogenesis involve distinct interconnected subpopulations: a combined computational and experimental study , ENeuro, 2 (2015)
work page 2015
-
[54]
A. K. Tryba, F. Pe ˜na, S. P. Lieske, J.-C. Viemari, M. Thoby-Brisson, and J.-M. Ramirez , Differential modulation of neural network and pacemaker activity underlying eupnea and sigh-breathing activities, Journal of neurophysiology, 99 (2008), pp. 2114–2125
work page 2008
-
[55]
J. Viemari, M. Bevengut, H. Burnet, P. Coulon, J.-M. Pequignot, M. Tiveron, and G. Hilaire , Phox2a gene, a6 neurons, and noradrenaline are essential for development of normal respiratory rhythm in mice , Journal of Neuroscience, 24 (2004), pp. 928–937
work page 2004
-
[56]
J.-C. Viemari and J.-M. Ramirez , Norepinephrine differentially modulates different types of respiratory pacemaker and nonpacemaker neurons, Journal of neurophysiology, 95 (2006), pp. 2070–2082
work page 2006
-
[57]
Y. W ang and J. E. Rubin , Multiple timescale mixed bursting dynamics in a respiratory neuron model , Journal of Computational Neuroscience, 41 (2016), pp. 245–268
work page 2016
-
[58]
Y. W ang and J. E. Rubin, Timescales and mechanisms of sigh-like bursting and spiking in models of rhythmic respiratory neurons, The Journal of Mathematical Neuroscience, 7 (2017), pp. 1–39
work page 2017
-
[59]
Y. W ang and J. E. Rubin , Complex bursting dynamics in an embryonic respiratory neuron model , Chaos: An Interdis- ciplinary Journal of Nonlinear Science, 30 (2020)
work page 2020
-
[60]
S. Zanella, A. Doi, A. J. Garcia, F. Elsen, S. Kirsch, A. D. Wei, and J.-M. Ramirez , When norepinephrine becomes a driver of breathing irregularities: how intermittent hypoxia fundamentally alters the modulatory response of the respiratory network, Journal of Neuroscience, 34 (2014), pp. 36–50
work page 2014
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.