Single-photon superradiance and subradiance in helical collectives of quantum emitters
Pith reviewed 2026-05-18 04:56 UTC · model grok-4.3
The pith
Continuous distributions of two-level systems on an infinite helix yield closed-form expressions for collective decay rates and Lamb shifts under single-photon excitation.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
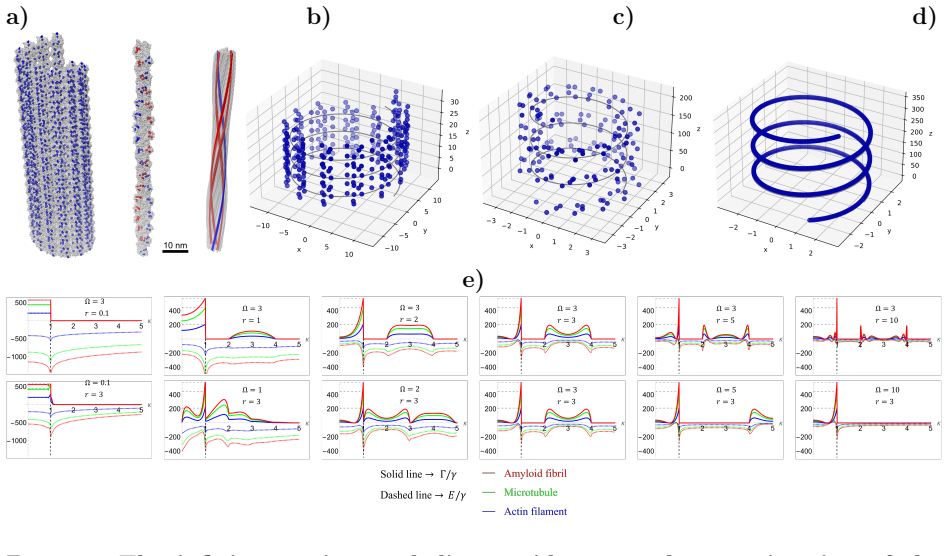
Novel analytical expressions are obtained for the collective decay rates and Lamb shifts of a single photon coupled to a continuous distribution of two-level systems placed along an infinite line and along an infinite helix. These expressions are compared with the eigenvalues for cylindrical arrangements, revealing dimensional limits in which the spectra coincide. Direct comparison with discrete lattices shows that the continuous scalar model does not recover the discrete vector results even in the zero-spacing limit, and that short-, intermediate-, and long-range interaction terms produce quantitatively different spectra. The helix formulas are then used to estimate the maximally superradiy
What carries the argument
The closed-form eigenvalue spectrum of the continuous interaction kernel for an infinite helical chain of two-level systems.
If this is right
- The fraction of trapped states and the thermally averaged decay rate for helical protein-fiber architectures can be read off directly from the analytical spectrum without large-scale numerics.
- Limits exist in which the collective eigenvalues of line, helix, and cylinder geometries become identical, allowing dimensional reduction in device design.
- Inclusion or exclusion of the near-field 1/r^3 term changes the spectrum qualitatively, so device engineering must retain the full interaction if accuracy at short range is required.
- Sparse emitter arrangements in helical geometries are predicted to exhibit excellent agreement between the continuous formulas and discrete simulations, guiding experimental densities.
Where Pith is reading between the lines
- The same continuous-kernel technique could be applied to other periodic or quasi-periodic curves, such as toroidal or helical bundles, to generate further closed-form spectra.
- For dense arrangements the persistent discrepancy between continuous scalar and discrete vector models suggests that hybrid models retaining discreteness at short scales may be needed.
- If the analytical estimates hold for real biomolecular fibers, they supply a parameter-free starting point for designing subradiant quantum memories or superradiant error-corrected channels inside living matter.
Load-bearing premise
The emitters can be modeled as a continuous distribution rather than discrete points, and the infinite-helix geometry remains a valid approximation for the finite, discrete arrangements found in real protein fibers.
What would settle it
Numerical diagonalization of the interaction matrix for a finite but long discrete helix whose spacing and density match a protein fiber, followed by comparison of the resulting decay-rate distribution against the analytical helix formulas, would falsify the continuous approximation if the mismatch exceeds the reported sparse-arrangement agreement.
Figures





read the original abstract
Collective emission of light from distributions of two-level systems (TLSs) was first predicted in 1954 by Robert Dicke, who showed that when $N$ quantum emitters absorb photons, their collective radiative decay rate can be enhanced (superradiance) or suppressed (subradiance) relative to a single emitter. In this work, we derive novel analytical expressions for the collective decay rates and Lamb shifts for the interaction of a single photon with a continuous distribution of TLSs on an infinite line and an infinite helix. We compare these solutions to collectives of TLSs on a cylinder, finding limits in which the eigenvalues of structures of different dimensions are equal. We also compare our solution with arrangements where the emitter distribution is discrete rather than continuous, and when short- ($1/r^3$), intermediate- ($1/r^2$), and long-range ($1/r$) interaction terms are included. We find important differences between the discrete vector and continuous scalar emitter cases, which do not agree in the limit where discrete spacing goes to 0. The analytical solution for the helix is then used to make estimates of the maximally superradiant state, thermally averaged collective decay rate, and percentage of trapped states of quantum emitter architectures in protein fibers. Given the differences between our idealized infinite helix and the numerical model describing protein fibers, our analytical estimates show excellent agreement with the numerical results for sparse arrangements of emitters in protein fibers. Our work thus bridges the gap between different formalisms for superradiance, aids the engineering of devices which harness quantum optical effects for computing with superradiant error correction and subradiant memories, and motivates the discovery and creation of flexible platforms for quantum information processing using the intrinsic helical geometries of biomatter.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper derives novel analytical expressions for the collective decay rates and Lamb shifts for the interaction of a single photon with a continuous distribution of TLSs on an infinite line and an infinite helix. It compares these solutions to collectives of TLSs on a cylinder, to discrete emitter arrangements, and across short-, intermediate-, and long-range interaction terms. The work identifies important differences between the continuous scalar and discrete vector cases that persist as spacing approaches zero, then applies the helix solution to estimate the maximally superradiant state, thermally averaged collective decay rate, and fraction of trapped states for quantum emitter architectures in protein fibers, reporting excellent agreement for sparse arrangements.
Significance. If the derivations and comparisons hold, the manuscript supplies closed-form analytical tools for superradiance and subradiance in helical geometries that bridge continuous and discrete formalisms. The explicit construction of parameter-free expressions from first principles for the infinite helix and line cases, together with the direct comparison to protein-fiber numerics, constitutes a concrete strength that could support engineering of superradiant error-correction schemes and subradiant memories.
major comments (1)
- [Application to protein fibers (comparison of analytical helix to numerical model)] The central application to protein fibers rests on the claim that the infinite continuous scalar helix provides useful estimates despite the noted differences from discrete vector models. The abstract states that the two cases 'do not agree in the limit where discrete spacing goes to 0' yet 'show excellent agreement with the numerical results for sparse arrangements.' Please identify the specific section or figure that quantifies the residual discrepancy in the sparse regime relevant to protein fibers and state the criterion used to judge the approximation adequate for the reported estimates of superradiant and trapped states.
minor comments (1)
- [Comparison with discrete arrangements] Clarify the precise definition of 'continuous distribution' versus 'discrete spacing approaching zero' when the interaction kernel includes the full 1/r^3, 1/r^2, and 1/r terms; a short paragraph or footnote would remove ambiguity for readers comparing to prior discrete-dipole literature.
Simulated Author's Rebuttal
We thank the referee for their careful reading of the manuscript and for identifying an opportunity to strengthen the presentation of our comparison between the analytical helix model and the numerical protein-fiber results. We address the major comment below and will incorporate the requested clarifications in the revised version.
read point-by-point responses
-
Referee: The central application to protein fibers rests on the claim that the infinite continuous scalar helix provides useful estimates despite the noted differences from discrete vector models. The abstract states that the two cases 'do not agree in the limit where discrete spacing goes to 0' yet 'show excellent agreement with the numerical results for sparse arrangements.' Please identify the specific section or figure that quantifies the residual discrepancy in the sparse regime relevant to protein fibers and state the criterion used to judge the approximation adequate for the reported estimates of superradiant and trapped states.
Authors: We agree that an explicit quantification of the residual discrepancy would improve clarity. The relevant comparisons appear in Section 5, with the superradiant decay rates, thermally averaged rates, and trapped-state fractions shown in Figures 8 and 9 for both the analytical helix and the discrete numerical model of the protein fiber. These figures demonstrate that the analytical predictions track the numerical results closely for inter-emitter spacings greater than approximately 0.1 wavelengths. However, the manuscript does not currently include a dedicated error metric or residual plot. In the revised manuscript we will add a new panel (or supplementary figure) that explicitly plots the relative difference (in percent) between the analytical and numerical values of the maximally superradiant rate and the trapped-state fraction as a function of spacing in the sparse regime. We will define the adequacy criterion as agreement to within 10% relative error for the key observables; this threshold is chosen because it preserves the correct ordering of superradiant versus subradiant behavior and provides useful order-of-magnitude guidance for biomolecular architectures. We view this as a minor but worthwhile clarification rather than a change to the scientific conclusions. revision: yes
Circularity Check
Analytical derivations for continuous helix and line are self-contained first-principles results
full rationale
The paper derives novel closed-form expressions for collective decay rates and Lamb shifts directly from the single-photon interaction Hamiltonian with continuous scalar TLS distributions on infinite line and helix geometries. These expressions are obtained by solving the eigenvalue problem for the interaction kernel in the continuous limit and are then compared (not fitted) to discrete vectorial cases, cylinder geometries, and varying interaction ranges. No load-bearing step reduces to a fitted parameter renamed as prediction, a self-citation chain, or an ansatz smuggled from prior work by the same authors; the protein-fiber estimates are presented as applications of the derived formulas with explicit caveats on the continuous-vs-discrete and infinite-vs-finite discrepancies. The derivation chain therefore remains independent of its target outputs.
Axiom & Free-Parameter Ledger
axioms (1)
- standard math Two-level systems (TLSs) interact via electromagnetic fields with short-range (1/r^3), intermediate (1/r^2), and long-range (1/r) terms.
Lean theorems connected to this paper
-
IndisputableMonolith/Foundation/AbsoluteFloorClosure.leanabsolute_floor_iff_bare_distinguishability unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
We derive novel analytical expressions for the collective decay rates and Lamb shifts for the interaction of a single photon with a continuous distribution of TLSs on an infinite line and an infinite helix... We find important differences between the discrete vector and continuous scalar emitter cases, which do not agree in the limit where discrete spacing goes to 0.
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
R. H. Dicke. Coherence in Spontaneous Radiation Processes.Phys. Rev., 93:99–110, Jan 1954
work page 1954
-
[2]
A. Asenjo-Garcia, M. Moreno-Cardoner, A. Albrecht, H. J. Kimble, and D. E. Chang. Exponential Improvement in Photon Storage Fidelities Using Subradiance and “Selec- tive Radiance” in Atomic Arrays.Phys. Rev. X, 7(3):031024, August 2017
work page 2017
-
[3]
Hamza Patwa, Nathan S. Babcock, and Philip Kurian. Quantum-enhanced photopro- tection in neuroprotein architectures emerges from collective light-matter interactions. Front. Phys., 12, aug 2024
work page 2024
-
[4]
N. S. Babcock, G. Montes-Cabrera, K. E. Oberhofer, M. Chergui, G. L. Celardo, and P. Kurian. Ultraviolet Superradiance from Mega-Networks of Tryptophan in Biological Architectures.J. Phys. Chem. B, 128(17):4035–4046, April 2024. 25
work page 2024
-
[5]
Svidzinsky, Fu Li, Hongyuan Li, Xiwen Zhang, C
Anatoly A. Svidzinsky, Fu Li, Hongyuan Li, Xiwen Zhang, C. H. Raymond Ooi, and Marlan O. Scully. Single-photon superradiance and radiation trapping by atomic shells. Phys. Rev. A, 93(4):043830, April 2016
work page 2016
-
[6]
On the existence of superradiant excitonic states in microtubules.New J
G L Celardo, M Angeli, T J A Craddock, and P Kurian. On the existence of superradiant excitonic states in microtubules.New J. Phys., 21(2):023005, feb 2019
work page 2019
-
[7]
Efficient light harvesting and photon sensing via engineered cooperative effects
Francesco Mattiotti, Mohan Sarovar, Giulio G Giusteri, Fausto Borgonovi, and G Luca Celardo. Efficient light harvesting and photon sensing via engineered cooperative effects. New J. Phys., 24(1):013027, 2022
work page 2022
-
[8]
Patrik R. Callis. [7] 1La and 1Lb transitions of tryptophan: Applications of theory and experimental observations to fluorescence of proteins. InFlourescence Spectroscopy, volume 278 ofMethods Enzymol., pages 113–150. Academic Press, 1997
work page 1997
-
[9]
S. Schenkl, F. van Mourik, G. van der Zwan, S. Haacke, and M. Chergui. Probing the Ultrafast Charge Translocation of Photoexcited Retinal in Bacteriorhodopsin.Science, 309(5736):917–920, August 2005
work page 2005
-
[10]
Refined structure ofα β-tubulin at 3.5 ˚A resolution.J
Jan L¨ owe, H Li, KH Downing, and E Nogales. Refined structure ofα β-tubulin at 3.5 ˚A resolution.J. Mol. Biol., 313(5):1045–1057, 2001
work page 2001
-
[11]
Falk Liberta, Sarah Loerch, Matthies Rennegarbe, Angelika Schierhorn, Per West- ermark, Gunilla T Westermark, Bouke PC Hazenberg, Nikolaus Grigorieff, Marcus F¨ andrich, and Matthias Schmidt. Cryo-EM fibril structures from systemic AA amy- loidosis reveal the species complementarity of pathological amyloids.Nat. Comm., 10(1):1–10, 2019
work page 2019
-
[12]
Pinar S Gurel, Laura Y Kim, Paul V Ruijgrok, Tosan Omabegho, Zev Bryant, and Gregory M Alushin. Cryo-EM structures reveal specialization at the myosin VI-actin interface and a mechanism of force sensitivity.Elife, 6:e31125, 2017
work page 2017
-
[13]
Suyash Bajpai, Masashi Aono, and Philip Kurian. Tracking and Distinguishing Slime Mold Solutions to the Traveling Salesman Problem through Synchronized Amplification in the Non-Equilibrium Steady State.arXiv:2504.03492, 2025
-
[14]
H. H. Jen. Phase-imprinted multiphoton subradiant states.Phys. Rev. A, 96(2):023814, August 2017
work page 2017
-
[15]
Marlan O. Scully. Collective Lamb Shift in Single Photon Dicke Superradiance.Phys. Rev. Lett., 102(14):143601, April 2009
work page 2009
-
[16]
Anatoly A. Svidzinsky and Marlan O. Scully. Evolution of collective N atom states in single photon superradiance: Effect of virtual Lamb shift processes.Opt. Commun., 282(14):2894–2897, July 2009
work page 2009
-
[17]
Marlan O. Scully. Single Photon Subradiance: Quantum Control of Spontaneous Emis- sion and Ultrafast Readout.Phys. Rev. Lett., 115(24):243602, December 2015. 26
work page 2015
-
[18]
Chengdong Mi, Khan Sadiq Nawaz, Liangchao Chen, Pengjun Wang, Han Cai, Da-Wei Wang, Shi-Yao Zhu, and Jing Zhang. Time-resolved interplay between superradiant and subradiant states in superradiance lattices of Bose-Einstein condensates.Phys. Rev. A, 104(4):043326, October 2021
work page 2021
-
[19]
Peter, Stefan Ostermann, and Susanne F
Jonah S. Peter, Stefan Ostermann, and Susanne F. Yelin. Chirality dependent photon transport and helical superradiance.Phys. Rev. Res., 6(2):023200, May 2024
work page 2024
-
[20]
Computational capacity of life in relation to the universe.Sci
Philip Kurian. Computational capacity of life in relation to the universe.Sci. Adv., 11(13):eadt4623, March 2025
work page 2025
-
[21]
Direct detection of a single photon by humans.Nat
Jonathan N Tinsley, Maxim I Molodtsov, Robert Prevedel, David Wartmann, Jofre Espigule-Pons, Mattias Lauwers, and Alipasha Vaziri. Direct detection of a single photon by humans.Nat. Commun., 7(1):12172, July 2016
work page 2016
-
[22]
S Hecht, S Shlaer, and M H Pirenne. Energy, quanta, and vision.J. Gen. Physiol., 25(6):819–840, July 1942
work page 1942
-
[23]
Retinal noise and absolute threshold.J
H B Barlow. Retinal noise and absolute threshold.J. Opt. Soc. Am., 46(8):634–639, August 1956
work page 1956
-
[24]
Haruhisa Okawa and Alapakkam P Sampath. Optimization of single-photon response transmission at the rod-to-rod bipolar synapse.Physiology (Bethesda), 22(4):279–286, August 2007
work page 2007
-
[25]
Retinal processing near absolute threshold: from behavior to mechanism.Annu
Greg D Field, Alapakkam P Sampath, and Fred Rieke. Retinal processing near absolute threshold: from behavior to mechanism.Annu. Rev. Physiol., 67(1):491–514, 2005
work page 2005
-
[26]
Possible entanglement detection with the naked eye.Phys
Nicolas Brunner, Cyril Branciard, and Nicolas Gisin. Possible entanglement detection with the naked eye.Phys. Rev. A, 78(5):052110, November 2008
work page 2008
-
[27]
Edge States and Topological Invariants of Non-Hermitian Systems.Phys
Shunyu Yao and Zhong Wang. Edge States and Topological Invariants of Non-Hermitian Systems.Phys. Rev. Lett., 121(8):086803, August 2018
work page 2018
-
[28]
Non-Hermitian Topological Invariants in Real Space.Phys
Fei Song, Shunyu Yao, and Zhong Wang. Non-Hermitian Topological Invariants in Real Space.Phys. Rev. Lett., 123(24):246801, December 2019. 27
work page 2019
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.