Seeing Below the Limit of Detection: A Censored-Poisson Bayesian Latent-Growth Change-Point Detector (the Span Detector) for Serial ctDNA in HR+/HER2- Metastatic Breast Cancer
Pith reviewed 2026-06-27 07:52 UTC · model grok-4.3
The pith
Span treats non-detects as censored observations to double early detection of indolent breast cancer progression from serial ctDNA.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
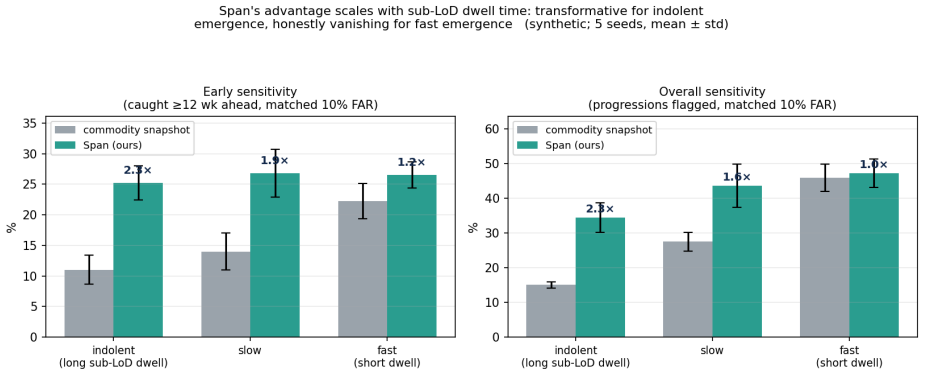
Span is a censored-Poisson Bayesian latent-growth change-point detector that models the binary detection process, accumulates a sequential generalised-likelihood-ratio statistic for an upward change-point in the per-variant detection rate, and raises competing-risks alarms with calibrated false-alarm control. On a synthetic cohort of HR+/HER2- metastatic breast cancer on CDK4/6-inhibitor plus endocrine therapy, at a matched 10% false-alarm rate, Span roughly doubles the fraction of impending progressions caught three months ahead in the indolent regime (25% vs 11% for the snapshot), with a falsifiable dose-response that is large for indolent emergence and vanishes for fast emergence. A value
What carries the argument
The Span detector, a censored-Poisson Bayesian latent-growth change-point model that accumulates evidence from left-censored non-detects to identify upward shifts in per-variant detection rate.
If this is right
- The performance advantage appears only in indolent regimes where detection is intermittent and disappears for rapid progression.
- Modeling detection patterns rather than measured values drives the gain, since a value-trajectory baseline matches the snapshot method.
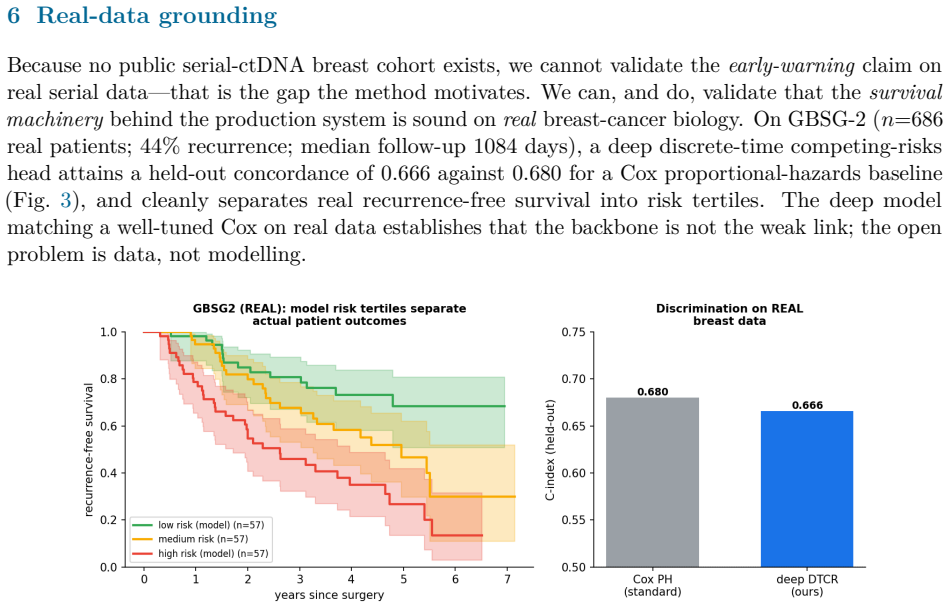
- The pipeline matches a Cox baseline on real survival data and correctly withholds improvement on a clean biomarker cohort, confirming regime specificity.
- Absence of learned weights eliminates overfitting as an explanation for the reported results.
Where Pith is reading between the lines
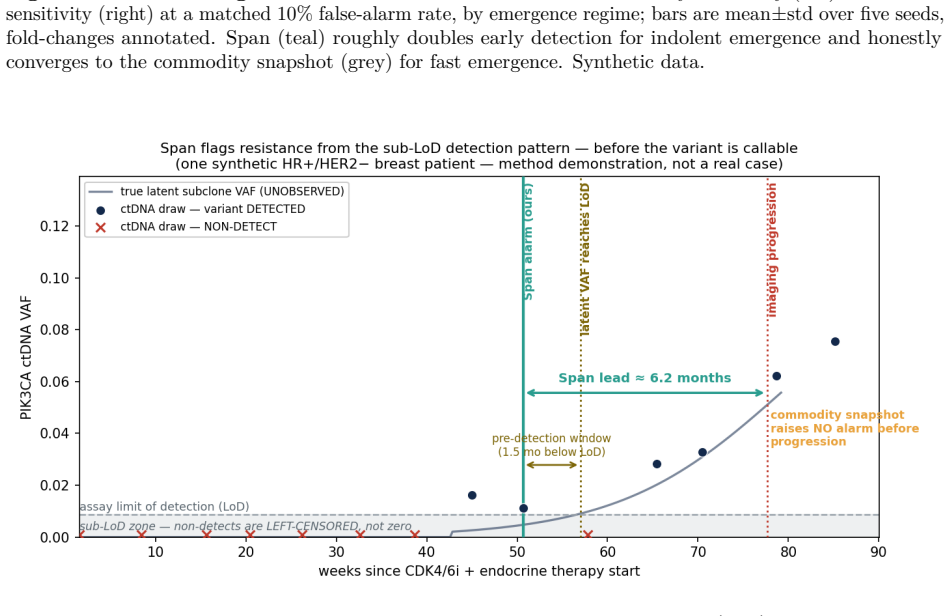
- If the synthetic trajectories match real detection intermittency, the same censored modeling could be tested on other liquid-biopsy settings with sub-LoD signals.
- The dose-response pattern suggests stratifying future real-data trials by estimated progression speed to isolate where the method adds value.
- Integration into monitoring would require prospective calibration of the false-alarm threshold against clinical action thresholds.
Load-bearing premise
The synthetic ctDNA trajectories accurately reproduce the intermittent detection statistics of real nascent subclones below the assay limit of detection.
What would settle it
A head-to-head comparison of Span versus snapshot methods on a real longitudinal ctDNA dataset from metastatic breast cancer patients, testing whether the doubling of three-month-ahead warnings at 10% false-alarm rate holds specifically for indolent progression cases.
Figures
read the original abstract
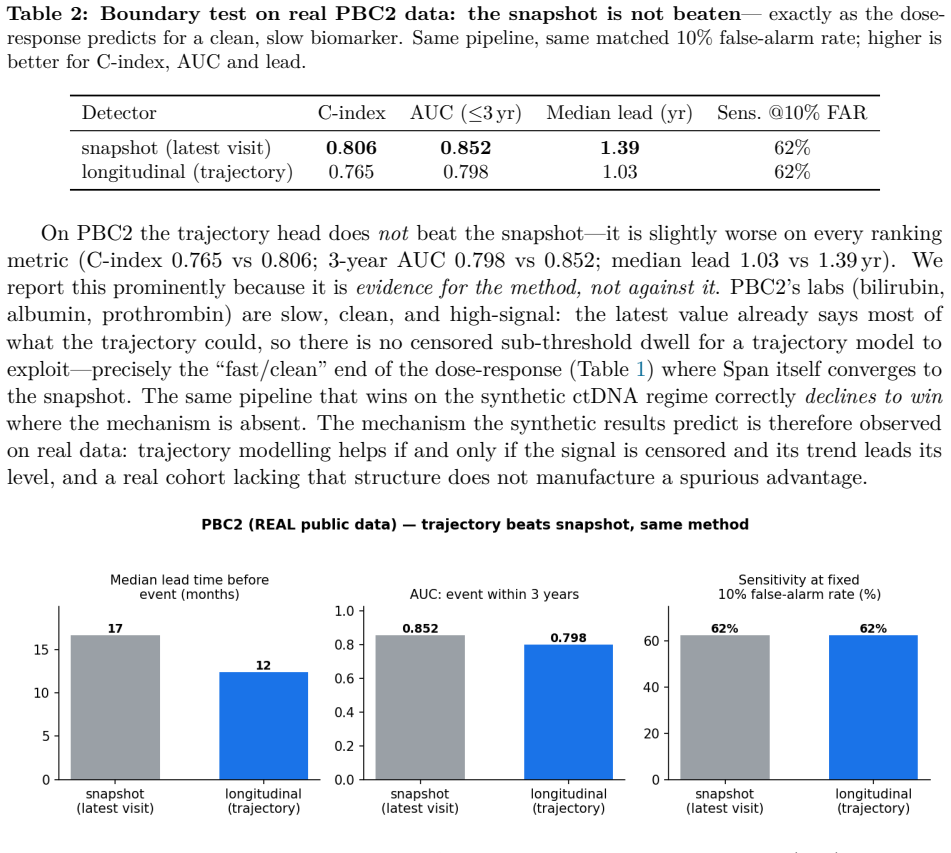
Circulating-tumour DNA (ctDNA) carries evidence of drug resistance months before imaging shows it, but the earliest evidence lives below the assay's limit of detection (LoD): a nascent subclone is detected only intermittently, producing a flickering sequence of faint detects and non-detects. Commercial liquid biopsies treat each draw as an independent snapshot and a non-detect as nothing. We argue a non-detect is a left-censored observation, and the pattern of non-detects and faint detects over time carries actionable evidence of growth before any single value is trustworthy. We introduce Span, a censored-Poisson Bayesian latent-growth change-point detector that models the binary detection process, accumulates a sequential generalised-likelihood-ratio statistic for an upward change-point in the per-variant detection rate, and raises a competing-risks alarm with calibrated false-alarm control. Span has no learned weights, so there is nothing to overfit. On a synthetic cohort of HR+/HER2- metastatic breast cancer on first-line CDK4/6-inhibitor plus endocrine therapy, at a matched 10% false-alarm rate, Span roughly doubles the fraction of impending progressions caught three months ahead (indolent regime: 25% vs 11% for the snapshot), with a falsifiable dose-response: large for indolent emergence, vanishing for fast emergence. A value-trajectory baseline performs identically to the snapshot, isolating the gain to the censored detection model. The survival backbone matches a Cox baseline on real breast-cancer data (GBSG-2, n=686; C-index 0.67 vs 0.68), and on a real longitudinal cohort with clean biomarkers (PBC2, n=312) the same pipeline correctly declines to win, a falsifiable boundary test confirming the mechanism is regime-specific. All ctDNA trajectories are synthetic.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper introduces Span, a parameter-free censored-Poisson Bayesian latent-growth change-point detector for serial ctDNA monitoring in HR+/HER2- metastatic breast cancer. It treats non-detects as left-censored observations, accumulates a sequential GLR statistic on the per-variant detection rate, and issues competing-risks alarms with calibrated false-alarm control. On synthetic ctDNA trajectories, at matched 10% FAR, Span doubles the fraction of impending progressions detected three months ahead in the indolent regime (25% vs 11% for snapshot baseline); the gain vanishes for fast emergence. Real-data experiments on GBSG-2 (Cox C-index match) and PBC2 (correct non-improvement) serve as boundary tests. All ctDNA trajectories are synthetic.
Significance. If the synthetic generator's fidelity to real below-LoD intermittency holds, Span offers a practical, non-overfit method to extract earlier resistance signals from existing liquid-biopsy assays. The parameter-free construction, explicit falsifiable dose-response, and use of real cohorts as negative controls are methodological strengths that distinguish it from learned models. The work could influence serial ctDNA interpretation protocols if the synthetic-to-real gap is closed.
major comments (2)
- [Abstract] Abstract (final sentence) and § on synthetic cohort: the central performance claim (25% vs 11% detection at 3 months, indolent regime) rests exclusively on synthetic trajectories; the manuscript provides no real longitudinal ctDNA series containing verified nascent subclones below LoD to calibrate or validate the generator's detection-rate process, censoring statistics, or growth-change parameters. This assumption is load-bearing for the dose-response claim.
- [Real-data experiments] Real-data experiments section: GBSG-2 (n=686) and PBC2 (n=312) test only the survival-backbone Cox component and correctly show no spurious gain; they do not validate the censored-Poisson change-point mechanism on ctDNA, leaving the ctDNA-specific performance unanchored to real intermittent detection behavior.
minor comments (2)
- [Methods] Clarify in the methods how the synthetic generator's parameters (e.g., detection probability below LoD, growth rates) were chosen and whether sensitivity analyses were performed.
- [Figures] Figure legends should explicitly state that all ctDNA panels are synthetic and note the exact FAR matching procedure used for the snapshot baseline.
Simulated Author's Rebuttal
We thank the referee for the detailed and constructive report. We agree that the central performance claims rest on synthetic ctDNA trajectories and that the real-data experiments do not validate the change-point mechanism itself. We respond to each major comment below and indicate the revisions we will make.
read point-by-point responses
-
Referee: [Abstract] Abstract (final sentence) and § on synthetic cohort: the central performance claim (25% vs 11% detection at 3 months, indolent regime) rests exclusively on synthetic trajectories; the manuscript provides no real longitudinal ctDNA series containing verified nascent subclones below LoD to calibrate or validate the generator's detection-rate process, censoring statistics, or growth-change parameters. This assumption is load-bearing for the dose-response claim.
Authors: We acknowledge that the performance numbers are generated exclusively from synthetic trajectories, as the manuscript already states. No real longitudinal ctDNA series with verified nascent subclones below LoD is used for calibration or validation of the generator. We will revise the abstract's final sentence to explicitly qualify the 25% vs 11% claim as synthetic and add a dedicated paragraph in the discussion that details the generator's modeling assumptions, the falsifiable dose-response property, and the current inability to anchor it to real below-LoD intermittency. revision: yes
-
Referee: [Real-data experiments] Real-data experiments section: GBSG-2 (n=686) and PBC2 (n=312) test only the survival-backbone Cox component and correctly show no spurious gain; they do not validate the censored-Poisson change-point mechanism on ctDNA, leaving the ctDNA-specific performance unanchored to real intermittent detection behavior.
Authors: This is correct. The GBSG-2 and PBC2 experiments are designed solely as boundary tests for the survival component and to demonstrate regime-specificity (no spurious improvement where none is expected). They do not claim to validate the censored-Poisson change-point detector on ctDNA. We will revise the section heading and text to label these experiments explicitly as negative controls for the non-ctDNA pipeline elements and to restate that ctDNA-specific evaluation remains synthetic. revision: yes
- No real longitudinal ctDNA series containing verified nascent subclones below the limit of detection is available to the authors, so direct empirical validation or calibration of the synthetic generator's detection-rate and censoring process cannot be performed in the current work.
Circularity Check
No significant circularity; main claim is empirical performance on independently generated synthetic trajectories
full rationale
The paper's derivation chain consists of a parameter-free censored-Poisson change-point detector applied to a synthetic ctDNA cohort, with the headline performance (25% vs 11% detection at 3 months) obtained directly from that application. Real-data experiments (GBSG-2 Cox match, PBC2 non-improvement) are presented only as boundary tests confirming regime specificity, not as inputs that define the ctDNA result. No self-citations, fitted parameters renamed as predictions, or self-definitional steps appear in the provided text; the model explicitly has no learned weights. The synthetic generator's fidelity to real below-LoD statistics is an external validity question, not a reduction of the reported result to its own inputs by construction.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption Detection events follow a censored-Poisson process with latent growth rate that can exhibit an upward change-point
- ad hoc to paper Synthetic ctDNA trajectories accurately capture the statistical properties of real patient data below the limit of detection
Reference graph
Works this paper leans on
-
[1]
J. O. Brett et al. ESR1 mutation as an emerging clinical biomarker in metastatic HR+ breast cancer. Breast Cancer Research, 23:115, 2021
2021
-
[2]
Bidard et al
F.-C. Bidard et al. Switch to fulvestrant and palbociclib versus no switch in advanced breast cancer with rising ESR1 mutation (PADA-1).Lancet Oncology, 23(11):1367–1377, 2022
2022
-
[3]
Abbosh, A
C. Abbosh, A. M. Frankell et al. Tracking early lung-cancer evolution and ctDNA dynamics (TRACERx). Nature, 616:553–562, 2023
2023
-
[4]
C. Lee, W. R. Zame, J. Yoon, M. van der Schaar. DeepHit: a deep learning approach to survival analysis with competing risks.AAAI, 2018
2018
-
[5]
E. S. Page. Continuous inspection schemes.Biometrika, 41(1/2):100–115, 1954
1954
-
[6]
G. Lorden. Procedures for reacting to a change in distribution.Annals of Mathematical Statistics, 42(6):1897–1908, 1971
1908
-
[7]
J. Tobin. Estimation of relationships for limited dependent variables.Econometrica, 26(1):24–36, 1958
1958
-
[8]
Sarkar and A
A. Sarkar and A. Singh Thakur. OncoTraj v1: limits of single-snapshot tissue NGS for resistance prediction in EGFR-mutant NSCLC. Span AI technical report, 2026. 9
2026
-
[9]
Schumacher et al
M. Schumacher et al. Randomized2 ×2trial evaluating hormonal treatment and the duration of chemotherapy in node-positive breast cancer (GBSG-2).Journal of Clinical Oncology, 12(10):2086–2093, 1994. 10
2086
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.