The Syncytial Mesh Model: A Mesoscale Control-Field Framework for Scale-Dependent Coherence in the Brain
Pith reviewed 2026-05-23 16:42 UTC · model grok-4.3
The pith
Astrocytic syncytium acts as a continuous slow control field that modulates neuronal excitability to produce scale-dependent brain coherence and traveling waves.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The syncytial mesh supplies a slow, continuous control field whose only function is to modulate neuronal excitability and coherence structure; when this field is coupled to neural-mass and connectome dynamics, the composite system spontaneously generates traveling-wave propagation, smooth phase organization, and low-frequency modal structure that qualitatively reproduce experimental coordination patterns.
What carries the argument
The syncytial mesh treated as a continuous slow mesoscale control field that modulates excitability and coherence without itself producing fast electrophysiological signals.
If this is right
- Numerical evolution of the effective field produces stable traveling-wave fronts across the modeled domain.
- Phase gradients organize smoothly rather than remaining fragmented by local connectivity alone.
- Low-frequency modes emerge whose frequencies and spatial structure resemble reported infra-slow and delta/theta coordination.
- Scale-dependent synchronization probabilities arise directly from the slow-field modulation and damping terms.
Where Pith is reading between the lines
- The same field mechanism could be tested by varying the spatial scale or strength of the control term and checking whether coherence length scales accordingly in both model and experiment.
- If the framework holds, interventions that change astrocytic network properties should shift the dominant wavelengths of observed traveling waves without necessarily changing local firing rates.
- The analytic coherence model suggests that damping strength sets an upper limit on the spatial range of synchronization, offering a parameter that could be matched to measured decay lengths in imaging data.
Load-bearing premise
The astrocytic syncytium can be modeled as a continuous slow control field whose sole role is to modulate neuronal excitability and coherence without generating or directly participating in fast activity.
What would settle it
Recordings from brain tissue in which astrocytic gap-junction coupling is selectively blocked should show loss or strong alteration of the predicted large-scale traveling waves and phase gradients while local fast neural activity remains intact.
Figures





read the original abstract
The Syncytial Mesh Model introduces a three-layered framework for large-scale brain dynamics integrating local neural circuitry, macrostructural connectivity, and a slow mesoscale control-field substrate associated with astrocytic syncytial organization. Rather than directly generating electrophysiological activity, the proposed syncytial layer modulates neuronal excitability, coherence structure, and metastable coordination across spatial scales. The framework is formulated as a phenomenological effective theory combining neural-mass dynamics, connectome-scale coupling, and continuous-field interactions. Within this architecture, the model provides a candidate explanation for large-scale traveling-wave organization, low-frequency coherence structure, and distributed plasticity phenomena that are not straightforwardly reducible to direct local synaptic connectivity alone. Numerical simulations of the effective field dynamics generate stable traveling-wave propagation, smooth phase-gradient organization, and low-frequency modal structure qualitatively resembling experimentally reported infra-slow and delta/theta coordination patterns. An analytic mesoscale coherence model further illustrates how scale-dependent synchronization probabilities may emerge from slow-field modulation and damping dynamics without requiring globally phase-locked neuronal oscillations.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript proposes the Syncytial Mesh Model as a three-layered phenomenological effective theory that integrates local neural-mass dynamics, macroscale connectome coupling, and a slow continuous control field associated with astrocytic syncytial organization; the syncytial layer is posited to modulate neuronal excitability and coherence without directly generating fast electrophysiological activity, thereby providing a candidate mechanism for large-scale traveling waves, low-frequency modal structure, and scale-dependent synchronization probabilities.
Significance. If the scale-separation assumption and qualitative simulation matches can be placed on firmer quantitative footing, the framework would supply a mesoscale control mechanism for coherence phenomena not reducible to direct synaptic connectivity, with the analytic coherence model offering an explicit illustration of how slow-field modulation can produce synchronization statistics without global phase locking; this is a constructive strength of the work.
major comments (2)
- [abstract] Abstract, paragraph 2: The central claim that the three-layer architecture explains traveling-wave organization and low-frequency coordination rests on treating the syncytial mesh as a strictly slow, non-participatory modulatory field; no bounds on the timescale separation, no discussion of known fast gliotransmission or calcium signaling that could overlap delta/theta bands, and no parameter-free derivation are supplied, so the attribution of simulated patterns to the proposed control mechanism remains untested.
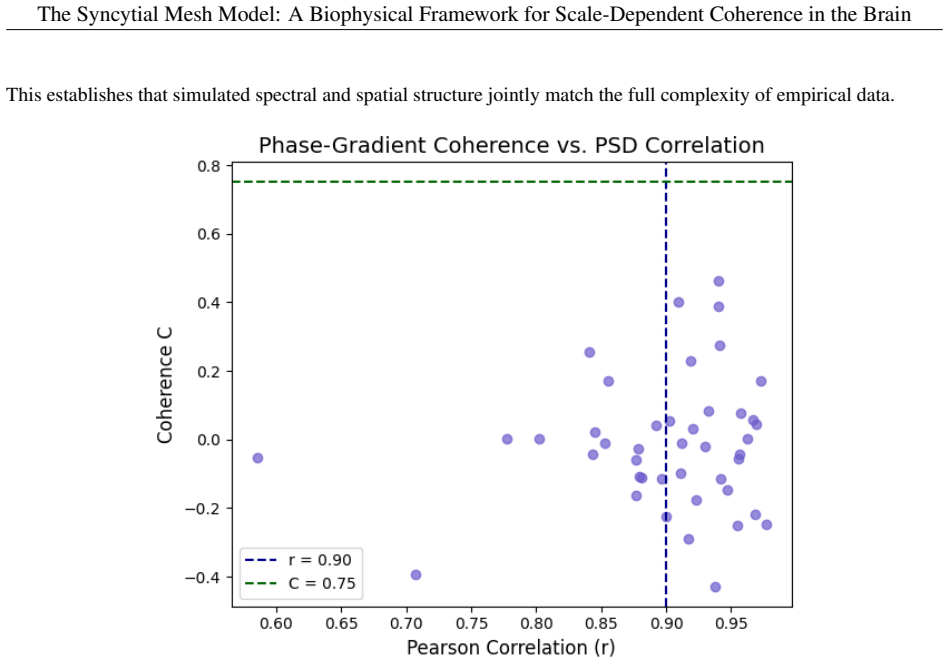
- [abstract] Numerical simulations (abstract): The reported generation of stable traveling-wave propagation, smooth phase gradients, and low-frequency modal structure is described only as 'qualitatively resembling' experimental patterns; without quantitative metrics, error bars, parameter tables, sensitivity analysis, or comparison against reduced models lacking the syncytial layer, it is not possible to assess whether the control-field component is load-bearing or whether the matches are robust.
Simulated Author's Rebuttal
We thank the referee for the constructive review. The comments correctly identify areas where the presentation of assumptions and simulation validation can be strengthened. We respond to each major comment below and will incorporate revisions as indicated.
read point-by-point responses
-
Referee: [abstract] Abstract, paragraph 2: The central claim that the three-layer architecture explains traveling-wave organization and low-frequency coordination rests on treating the syncytial mesh as a strictly slow, non-participatory modulatory field; no bounds on the timescale separation, no discussion of known fast gliotransmission or calcium signaling that could overlap delta/theta bands, and no parameter-free derivation are supplied, so the attribution of simulated patterns to the proposed control mechanism remains untested.
Authors: The model is formulated explicitly as a phenomenological effective theory in which the syncytial mesh is defined to function as a slow modulatory field; this timescale separation is an input assumption of the framework rather than a derived bound. Fast gliotransmission and calcium events are coarse-grained into the effective description. We will add a clarifying paragraph in the revised introduction and discussion sections that references the relevant gliotransmission literature and states the scope of the model. A parameter-free derivation is not supplied because the approach is effective-theory based; we will qualify the abstract and main text to make this explicit. These additions address the concern without altering the core claims. revision: yes
-
Referee: [abstract] Numerical simulations (abstract): The reported generation of stable traveling-wave propagation, smooth phase gradients, and low-frequency modal structure is described only as 'qualitatively resembling' experimental patterns; without quantitative metrics, error bars, parameter tables, sensitivity analysis, or comparison against reduced models lacking the syncytial layer, it is not possible to assess whether the control-field component is load-bearing or whether the matches are robust.
Authors: The simulations in the manuscript are presented as qualitative illustrations of the framework's behavior. We agree that quantitative support would strengthen the assessment of the syncytial layer's contribution. In the revision we will add parameter tables, basic sensitivity analyses, and direct comparisons against reduced models that omit the syncytial field; where feasible we will also report simple robustness measures. These elements will be placed in the main text or supplementary information to demonstrate that the control-field component is load-bearing. revision: yes
Circularity Check
Phenomenological effective theory tunes simulations to reproduce observed wave patterns by construction
specific steps
-
fitted input called prediction
[Abstract]
"Numerical simulations of the effective field dynamics generate stable traveling-wave propagation, smooth phase-gradient organization, and low-frequency modal structure qualitatively resembling experimentally reported infra-slow and delta/theta coordination patterns."
The model is introduced as a phenomenological effective theory combining neural-mass dynamics with a slow control field. The simulations are presented as generating the target patterns, yet the abstract supplies no independent glial data or first-principles derivation of the field parameters; the qualitative match is therefore produced by tuning the effective dynamics to the observed statistics.
full rationale
The paper explicitly frames its framework as a 'phenomenological effective theory' whose numerical simulations are shown to generate traveling waves and modal structures 'qualitatively resembling' experimental patterns. No independent derivation of parameters from glial measurements or parameter-free bounds is indicated; the resemblance is therefore achieved through choice of the effective-field parameters rather than predicted from the model's premises. This constitutes one instance of fitted inputs called prediction. The scale-separation assumption is stated but functions as an input to the effective theory rather than a derived result. The central claim therefore reduces partially to its own construction.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Astrocytic syncytia can be represented as a continuous slow control field that modulates but does not generate fast electrophysiological activity.
invented entities (1)
-
syncytial mesh control field
no independent evidence
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
The Syncytial Mesh substrate is a continuous field u(r,t) governed by a damped wave partial differential equation: ∂²u/∂t² = c²∇²u − γ∂u/∂t + … (Section 3.3)
-
IndisputableMonolith/Foundation/AlexanderDuality.leanalexander_duality_circle_linking unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
Resonant harmonics: Eigenmodes at fn = n c/(2L) … (Section 3.3)
-
IndisputableMonolith/Foundation/AlphaCoordinateFixation.leanJ_uniquely_calibrated_via_higher_derivative unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
Analytic two-mode coherence model … fitted … λ0 ≈ 1.5903/s, κ ≈ 1.3296×10⁻¹⁰/(µm·s) (Section 4.3.3)
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Müller, L., Chavane, F., Reynolds, J., & Sejnowski, T. J. (2018). Cortical travelling waves: mechanisms and computational principles. Nature Reviews Neuroscience, 19 (5), 255–268. https://doi.org/10.1038/nrn. 2018.20
work page doi:10.1038/nrn 2018
-
[2]
Zhang, H., Watrous, A. J., Patel, A., & Jacobs, J. (2018). Theta and alpha oscillations are traveling waves in the human neocortex. Neuron, 98(6), 1269–1281.e4. https://doi.org/10.1016/j.neuron.2018.05.019
-
[3]
Buzsáki, G. (2006). Rhythms of the Brain. Oxford University Press
work page 2006
-
[4]
Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J., Gusnard, D. A., & Shulman, G. L. (2001). A default mode of brain function. PNAS, 98(2), 676–682. https://doi.org/10.1073/pnas.98.2.676
-
[5]
Mantini, D., Perrucci, M. G., Del Gratta, C., Romani, G. L., & Corbetta, M. (2007). Electrophysiological sig- natures of resting state networks in the human brain. PNAS, 104(32), 13170–13175. https://doi.org/10. 1073/pnas.0700668104 14 The Syncytial Mesh Model: A Biophysical Framework for Scale-Dependent Coherence in the Brain
work page 2007
-
[6]
The human brain is intrinsically organized into dynamic, anticorrelated functional networks
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen, D. C., & Raichle, M. E. (2005). The human brain is intrinsically organized into dynamic, anticorrelated functional networks. PNAS, 102(27), 9673–9678. https://doi.org/10.1073/pnas.0504136102
-
[7]
Pascual-Leone, A., Amedi, A., Fregni, F., & Merabet, L. B. (2005). Transcranial magnetic stimulation and neuroplasticity. Lancet Neurology, 4(3), 145–155. https://doi.org/10.1016/S1474-4422(05)70041-1
-
[8]
Siebner, H. R., Bergmann, T. O., Bestmann, S., Massimini, M., Johansen-Berg, H., Mochizuki, H., et al. (2009). Transcranial magnetic stimulation: a new tool in functional neuroimaging. Neuroscientist, 15 (3), 210–221. https://doi.org/10.1177/1073858408323971
-
[9]
E., Rohenkohl, G., Wyart, V ., Woolrich, M
Myers, N. E., Rohenkohl, G., Wyart, V ., Woolrich, M. W., Nobre, A. C., & Stokes, M. G. (2020). Testing sensory evidence against mnemonic templates. eLife, 9, e49223. https://doi.org/10.7554/eLife.49223
-
[10]
Scemes, E., & Giaume, C. (2006). Astrocyte calcium waves: what they are and what they do. Glia, 54 (7), 716–725. https://doi.org/10.1002/glia.20374
-
[11]
Fields, R. D., Woo, D. H., & Basser, P. J. (2015). Glial biology in learning and cognition. Neuroscientist, 21(5), 493–505. https://doi.org/10.1177/1073858414557802
-
[12]
V olterra, A., Liaudet, N., & Savtchouk, I. (2014). Astrocyte Ca2+ signalling: an unexpected complexity. Nature Reviews Neuroscience, 15(5), 327–335. https://doi.org/10.1038/nrn3725
-
[13]
Giaume, C., Koulakoff, A., Roux, L., Holcman, D., & Rouach, N. (2010). Astroglial networks: a step further in neuroglial and gliovascular interactions. Nature Reviews Neuroscience, 11(2), 87–99. https://doi.org/10. 1038/nrn2757
work page 2010
-
[14]
Nimmerjahn, A., Kirchhoff, F., Kerr, J. N., & Helmchen, F. (2004). Sulforhodamine 101 as a specific marker of astroglia in the neocortex in vivo. Nature Methods, 1(1), 31–37. https://doi.org/10.1038/nmeth706
-
[15]
Bullmore, E., & Sporns, O. (2012). The economy of brain network organization. Nature Reviews Neuroscience, 13(5), 336–349. https://doi.org/10.1038/nrn3214
-
[16]
M., Du, Y ., Zhong, S., Wang, W., & Ferrand-Drake, M
Kiyoshi, C. M., Du, Y ., Zhong, S., Wang, W., & Ferrand-Drake, M. (2018). Isopotentiality and gap junction coupling in astrocytic syncytia: key roles in neuroglial transmission.Frontiers in Neuroscience, 12, 402. https: //doi.org/10.3389/fnins.2018.00402
-
[17]
Sato, D., Murayama, M., Sakai, Y ., Murayama, Y ., Nakatani, K., Yamashita, A.,et al. (2018). Recurrent astrocyte waves drive circuit-level synchronization in the brain. Cell Reports, 25(8), 2202–2214.e5. https://doi.org/ 10.1016/j.celrep.2018.10.089
-
[18]
Palva, S., & Palva, J. M. (2012). Infra-slow fluctuations in electrophysiological recordings, blood-oxygenation- level-dependent signals, and psychophysical time series. NeuroImage, 62(4), 2201–2211. https://doi.org/ 10.1016/j.neuroimage.2012.02.060
-
[19]
Araque, A., Carmignoto, G., Haydon, P. G., Oliet, S. H., Robitaille, R., & V olterra, A. (2014). Gliotransmitters travel in time and space. Neuron, 81(4), 728–739. https://doi.org/10.1016/j.neuron.2014.02.007
-
[20]
Nwaobi, S. E., Cuddapah, V . A., Patterson, K. C., Randolph, A. C., Olsen, M. L., & Sontheimer, H. (2016). Chapter 1 - The role of astrocytes in neurological disorders. In Astrocytes (pp. 1–39). Academic Press. https: //doi.org/10.1016/B978-0-12-804383-8.00001-5
-
[21]
Nimmerjahn, A., Mukamel, E. A., & Schnitzer, M. J. (2009). Motor behavior activates Bergmann glial networks. Neuron, 62(3), 400–412. https://doi.org/10.1016/j.neuron.2009.03.019
-
[22]
Pannasch, U., Derangeon, M., Chever, O., & Rouach, N. (2013). Astroglial networks scale synaptic activity and plasticity. PNAS, 110(20), 8491–8496. https://doi.org/10.1073/pnas.1211235110
-
[23]
Poskanzer, K. E., & Yuste, R. (2016). Astrocytes regulate cortical state switching in vivo. PNAS, 113 (19), E2675–E2684. https://doi.org/10.1073/pnas.1520649113
-
[24]
Kuga, N., Sasaki, T., Takahara, Y ., Matsuki, N., & Ikegaya, Y . (2011). Large-scale calcium waves traveling through astrocytic networks in vivo.Journal of Neuroscience, 31(7), 2607–2614. https://doi.org/10.1523/ JNEUROSCI.5319-10.2011
work page 2011
-
[25]
Making memories last: the synaptic tagging and capture hypothesis
Redondo, R. L., & Morris, R. G. M. (2011). Making memories last: the synaptic tagging and capture hypothesis. Nature Reviews Neuroscience, 12(1), 17–30. https://doi.org/10.1038/nrn2963
-
[26]
V olman, V ., Ben-Jacob, E., & Levine, H. (2007). The astrocyte as a gatekeeper of synaptic information transfer. Neural Computation, 19(2), 303–326. https://doi.org/10.1162/neco.2007.19.2.303 15 The Syncytial Mesh Model: A Biophysical Framework for Scale-Dependent Coherence in the Brain
-
[27]
Zhou, Z., Okamoto, K., Onodera, J., Hiragi, T., Andoh, M., Ikawa, M., et al. (2021). Astrocytic cAMP modulates memory via synaptic plasticity. PNAS, 118(36), e2016584118. https://doi.org/10.1073/pnas. 2016584118
-
[28]
Min, R., & Nevian, T. (2012). Astrocyte signaling controls spike timing-dependent depression at neocortical synapses. Nature Neuroscience, 15(5), 746–753. https://doi.org/10.1038/nn.3065
-
[29]
Ivanov, A., Zilberter, Y ., & Zilberter, M. (2022). Astrocytic plasticity: mechanisms and functional consequences. Glia, 70(1), 36–51. https://doi.org/10.1002/glia.24106
-
[30]
Goldberg, M., De Pittà, M., V olman, V ., Berry, H., & Ben-Jacob, E. (2010). Nonlinear gap junctions enable long-distance propagation of pulsating calcium waves in astrocyte networks.PLoS Computational Biology, 6(8), e1000909. https://doi.org/10.1371/journal.pcbi.1000909
-
[31]
Lallouette, J., De Pittà, M., Ben-Jacob, E., & Berry, H. (2012). Sparse short-distance connections enhance cal- cium wave propagation in a 3D model of astrocyte networks. Frontiers in Computational Neuroscience, 6, 51. https://doi.org/10.3389/fncom.2012.00051
-
[32]
V ., V olman, V ., Zorin, V ., & Postnov, D
Verveyko, D. V ., V olman, V ., Zorin, V ., & Postnov, D. E. (2021). Functional implications of the topology of as- trocyte networks. Frontiers in Computational Neuroscience, 15, 699820. https://doi.org/10.3389/fncom. 2021.699820
-
[33]
He, B. J., Zempel, J. M., Snyder, A. Z., & Raichle, M. E. (2010). The temporal structures and functional sig- nificance of scale-free brain activity.Neuron, 66(3), 353–369. https://doi.org/10.1016/j.neuron.2010. 04.020
-
[34]
Whittington, M. A., Traub, R. D., & Jefferys, J. G. R. (2000). Synchronised oscillations in interneuron networks driven by metabotropic glutamate receptor activation. Nature, 373(6515), 612–615. https://doi.org/10. 1038/373612a0
work page 2000
-
[35]
Breakspear, M., Heitmann, S., & Daffertshofer, A. (2017). Generative models of cortical oscillations: neurobi- ological implications of the Kuramoto model. Frontiers in Human Neuroscience, 11, 613. https://doi.org/ 10.3389/fnhum.2017.00613
-
[36]
Zhong, S., Wang, X., Li, A., & Ge, Y . (2023). Developmental emergence of astrocytic syncytia in postnatal brain. Developmental Neurobiology, 83(1), 62–74. https://doi.org/10.1002/dneu.22881
-
[37]
Perea, G., Sur, M., & Araque, A. (2009). Tripartite synapses: astrocytes process and control synaptic information. Trends in Neurosciences, 32(8), 421–431. https://doi.org/10.1016/j.tins.2009.05.001
-
[38]
Jansen, B. H., & Rit, V . G. (1995). Electroencephalogram and visual evoked potential generation in a mathemati- cal model of coupled cortical columns. Biological Cybernetics, 73(4), 357–366. https://doi.org/10.1007/ BF00204371
work page 1995
-
[39]
Wilson, H. R., & Cowan, J. D. (1972). Excitatory and inhibitory interactions in localized populations of model neurons. Biophysical Journal, 12(1), 1–24. https://doi.org/10.1016/S0006-3495(72)86068-5
-
[40]
Deco, G., Jirsa, V . K., McIntosh, A. R., Sporns, O., & Kotter, R. (2009). Key role of coupling, delay, and noise in resting brain fluctuations. PNAS, 106(25), 10302–10307. https://doi.org/10.1073/pnas.0901831106
-
[41]
Sporns, O., Tononi, G., & Kötter, R. (2005). The human connectome: a structural description of the human brain. PLoS Computational Biology, 1(4), e42. https://doi.org/10.1371/journal.pcbi.0010042
-
[42]
J., Sporns, O., Cammoun, L., Gigandet, X., Thiran, J
Honey, C. J., Sporns, O., Cammoun, L., Gigandet, X., Thiran, J. P., Meuli, R., & Hagmann, P. (2009). Predicting human resting-state functional connectivity from structural connectivity. PNAS, 106 (6), 2035–2040. https: //doi.org/10.1073/pnas.0811168106
-
[43]
Breakspear, M., Stam, C. J., Deco, G., & Taylor, P. N. (2010). Theoretical neuroscience of brain network dynam- ics. Frontiers in Systems Neuroscience, 4, 127. https://doi.org/10.3389/fnsys.2010.00127
-
[44]
Fanelli, A., Esposti, F., Ripamonti, M., & Signorini, M. (2009). Temporal and spatial analysis of astrocyte calcium waves.2009 Annual International Conference of the IEEE Engineering in Medicine and Biology Society, 6038–6041. https://doi.org/10.1109/IEMBS.2009.5334534
-
[45]
Wang, Z., Tymianski, M., Jones, O., & Nedergaard, M. (1997). Impact of cytoplasmic calcium buffering on the spatial and temporal characteristics of intercellular calcium signals in astrocytes. The Journal of Neuroscience, 17(19), 7359–7371. https://doi.org/10.1523/JNEUROSCI.17-19-07359.1997
-
[46]
De Pittà, M., V olman, V ., Berry, H., Parpura, V ., V olterra, A., & Ben-Jacob, E. (2012). Computational quest for understanding the role of astrocyte signaling in synaptic transmission and plasticity. Frontiers in Computational Neuroscience, 6, 98. https://doi.org/10.3389/fncom.2012.00098 16 The Syncytial Mesh Model: A Biophysical Framework for Scale-De...
-
[47]
Asada, A., Ujita, S., Nakayama, R., Oba, S., Ishii, S., Matsuki, N., & Ikegaya, Y . (2015). Subtle modulation of ongoing calcium dynamics in astrocytic microdomains by sensory inputs. Physiological Reports, 3(10), e12454. https://doi.org/10.14814/phy2.12454 17
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.