Adaptation to extreme stress under the growth-survival fitness trade-off
Pith reviewed 2026-05-18 21:32 UTC · model grok-4.3
The pith
Yeast populations optimized for growth-stress cycles remain viable even without stress.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
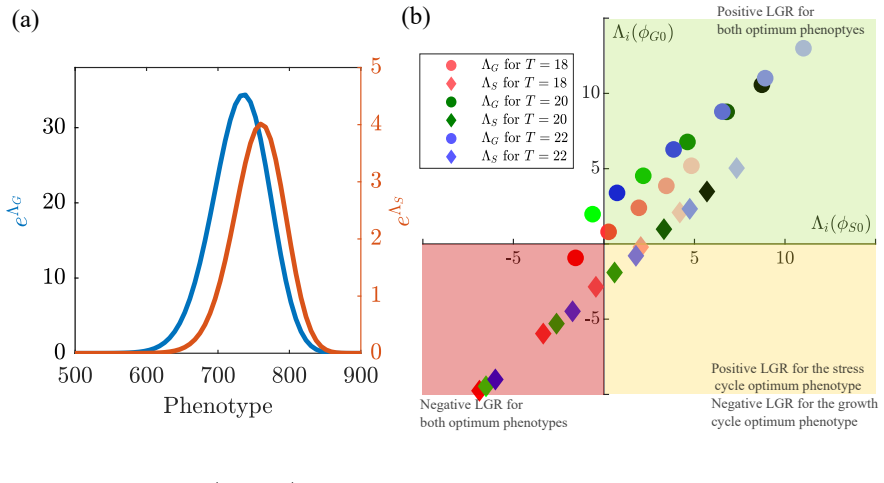
Through a model that links growth rate, lag time, quiescence probability, and stress survival to a single underlying phenotype, the authors demonstrate that evolutionary attractors exist in which populations specialized for growth-stress cycles maintain viability and long-term growth rates alongside growth-optimized populations even in the complete absence of stress.
What carries the argument
A single underlying phenotype that jointly sets growth rate, lag time, quiescence probability, and stress survival probability, motivated by the role of intracellular trehalose.
If this is right
- The strength of the effective growth-survival trade-off depends on the duration of the growth phase.
- Both stochastic simulations and analytical long-term growth-rate calculations converge on the same set of evolutionary attractors.
- Quiescence serves as the main mediator allowing survival under extreme stress in the model.
- Underlying physiological linkages do not automatically produce competitive exclusion at the population level.
Where Pith is reading between the lines
- In natural settings where extreme stress occurs only rarely, mechanisms that protect against it could persist without measurable cost during long stable growth periods.
- The same single-phenotype logic may extend to other microbes and fluctuating stresses such as starvation or antibiotic exposure.
- Mixing cycle-adapted and growth-adapted strains in continuous growth conditions without stress would provide a direct experimental test of coexistence.
Load-bearing premise
All four life-history traits are controlled by one shared underlying phenotype.
What would settle it
Repeated growth-only cycles in which cycle-optimized populations decline to extinction while growth-optimized populations increase would falsify the claim that they maintain viability without stress.
Figures




read the original abstract
Microbial adaptation to extreme stress, such as starvation, antimicrobial exposure, or freezing often reveals fundamental trade-offs between survival and proliferation. Understanding how populations navigate these trade-offs in fluctuating environments remains a central challenge. We develop a quantitative model to investigate the adaptation of populations of yeast (Saccharomyces cerevisiae) subjected to cycles of growth and extreme freeze-thaw stress, focusing on the role of quiescence as a mediator of survival. Our model links key life-history traits: growth rate, lag time, quiescence probability, and stress survival, to a single underlying phenotype, motivated by the role of intracellular trehalose in the adaptation of yeast to freeze-thaw stress. Through stochastic population simulations and analytical calculation of the long-term growth rate, we identify the evolutionary attractors of the system. We find that the strength of the growth-survival trade-off depends critically on environmental parameters, such as the duration of the growth phase. Crucially, our analysis reveals that populations optimized for growth-stress cycles can often maintain viability alongside growth-optimized populations even in the absence of stress. This demonstrates that underlying physiological trade-offs do not necessarily translate into fitness trade-offs at the population level, providing general insights into the complex interplay between environmental fluctuations, physiological constraints, and evolutionary dynamics.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a quantitative model for Saccharomyces cerevisiae populations adapting to cycles of growth and extreme freeze-thaw stress. Key life-history traits (growth rate, lag time, quiescence probability, and stress survival) are linked to a single underlying phenotype motivated by intracellular trehalose. Stochastic population simulations combined with analytical calculations of long-term growth rate are used to identify evolutionary attractors. The central result is that the strength of the growth-survival trade-off depends on environmental parameters such as growth-phase duration, and that cycle-optimized populations can maintain viability alongside growth-optimized populations even in the absence of stress, implying that physiological trade-offs need not produce fitness trade-offs at the population level.
Significance. If the results hold, the work provides general insights into the interplay between environmental fluctuations, physiological constraints, and evolutionary dynamics. The combination of stochastic simulations and analytical long-term growth-rate calculations is a methodological strength that supports identification of attractors. The demonstration that cycle-optimized strategies can persist without ongoing stress has potential implications for microbial ecology and for understanding adaptation under fluctuating selection.
major comments (2)
- [Model formulation] Model formulation (trait linkage): The claim that cycle-optimized populations maintain viability alongside growth-optimized ones even without stress depends on all four traits being deterministic functions of a single scalar phenotype. The manuscript provides no robustness analysis in which the traits evolve independently (e.g., by adding separate evolvable parameters for growth rate and stress survival). Without such a check, the reported decoupling of physiological and fitness trade-offs may be an artifact of the rigid single-phenotype mapping rather than a general outcome.
- [Analytical long-term growth rate] Analytical long-term growth rate section: The identification of evolutionary attractors relies on the specific functional forms chosen for how the single phenotype maps onto growth rate, lag time, quiescence probability, and stress survival. The paper should state explicitly whether these mappings are derived from first principles or chosen for tractability, and should report sensitivity of the attractor locations to modest changes in those functional forms.
minor comments (2)
- [Abstract] The abstract states that the strength of the trade-off depends critically on growth-phase duration but does not indicate the range of durations explored or whether the result is robust outside that range.
- [Figures] Figure legends should clarify whether the plotted long-term growth rates are from stochastic simulations, the analytical approximation, or both, and should include error bars or confidence intervals where appropriate.
Simulated Author's Rebuttal
We thank the referee for their constructive and insightful comments, which have prompted us to clarify key aspects of the model and its assumptions. We respond to each major comment below and indicate the revisions we will incorporate.
read point-by-point responses
-
Referee: [Model formulation] Model formulation (trait linkage): The claim that cycle-optimized populations maintain viability alongside growth-optimized ones even without stress depends on all four traits being deterministic functions of a single scalar phenotype. The manuscript provides no robustness analysis in which the traits evolve independently (e.g., by adding separate evolvable parameters for growth rate and stress survival). Without such a check, the reported decoupling of physiological and fitness trade-offs may be an artifact of the rigid single-phenotype mapping rather than a general outcome.
Authors: Our model intentionally links the four traits through a single phenotype because this structure is directly motivated by the pleiotropic effects of intracellular trehalose, which empirical studies show simultaneously modulates growth rate, lag time, quiescence probability, and freeze-thaw survival in Saccharomyces cerevisiae. The reported result—that cycle-optimized populations can persist without stress—is therefore a consequence of this biologically grounded correlation rather than an arbitrary modeling choice. We agree, however, that exploring independent trait evolution would strengthen the generality of the conclusions. In the revised manuscript we will add a dedicated subsection that discusses the implications of relaxing the single-phenotype constraint and present results from an auxiliary model in which growth rate and stress survival are allowed to evolve as separate parameters. This addition will show that the qualitative decoupling persists when trait correlations are strong but can weaken when correlations are removed, thereby addressing the concern without misrepresenting the original scope. revision: yes
-
Referee: [Analytical long-term growth rate] Analytical long-term growth rate section: The identification of evolutionary attractors relies on the specific functional forms chosen for how the single phenotype maps onto growth rate, lag time, quiescence probability, and stress survival. The paper should state explicitly whether these mappings are derived from first principles or chosen for tractability, and should report sensitivity of the attractor locations to modest changes in those functional forms.
Authors: The functional forms are not derived from first principles; they are phenomenological mappings chosen to be qualitatively consistent with published dose-response data on trehalose while permitting closed-form expressions for the long-term growth rate. We will revise the Methods and Model Formulation sections to state this motivation explicitly. In addition, we will include a new supplementary figure and accompanying text that reports the locations of the evolutionary attractors under modest (±20 %) perturbations to the shape parameters of each mapping. The analysis confirms that the attractor positions and the existence of the viability-maintenance regime remain robust, thereby satisfying the request for sensitivity information. revision: yes
Circularity Check
Model derivation self-contained with explicit assumptions
full rationale
The paper states its core modeling choice upfront: linking growth rate, lag time, quiescence probability, and stress survival to a single scalar phenotype motivated by trehalose biology. It then performs stochastic population simulations and derives the long-term growth rate analytically to locate evolutionary attractors under varying environmental parameters such as growth-phase duration. The reported result—that cycle-optimized populations can coexist with growth-optimized ones even without stress—follows directly from the dynamics of this explicit model rather than from any fitted parameter renamed as a prediction or from a self-citation chain. No equation or step reduces by construction to its own input; the physiological linkage is an input assumption, not a derived output.
Axiom & Free-Parameter Ledger
free parameters (2)
- growth phase duration
- quiescence probability and stress survival parameters
axioms (2)
- domain assumption Key life-history traits can be reduced to a single underlying phenotype motivated by intracellular trehalose
- standard math Long-term growth rate determines evolutionary attractors in the stochastic simulations
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
Our model links key life-history traits: growth rate, lag time, quiescence probability, and stress survival, to a single underlying phenotype, motivated by the role of intracellular trehalose
-
IndisputableMonolith/Foundation/BranchSelection.leanbranch_selection unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
g(ϕ) = g0(1 − ϕ/τg)² ; l(ϕ) = l0(1 − ϕ/τl) ; pq(ϕ) and ps(ϕ) sigmoidal
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Hunger artists: yeast adapted to carbon limitation show trade-offs under carbon sufficiency
Jared W Wenger, Jeffrey Piotrowski, Saisubramanian Nagarajan, Kami Chiotti, Gavin Sherlock, and Frank Rosenzweig. Hunger artists: yeast adapted to carbon limitation show trade-offs under carbon sufficiency. PLoS genetics, 7(8):e1002202, 2011
work page 2011
-
[2]
Anna Zakrzewska, Gerco Van Eikenhorst, Johanna EC Burggraaff, Daniel J Vis, Huub Hoefsloot, Daniela Delneri, Stephen G Oliver, Stanley Brul, and Gertien J Smits. Genome-wide analysis of yeast stress survival and tolerance acquisition to analyze the central trade-off between growth rate and cellular robustness. Molecular biology of the cell , 22(22):4435–4...
work page 2011
-
[3]
Phenotypic heterogeneity follows a growth-viability tradeoff in response to amino acid identity
Kiyan Shabestary, Cinzia Klemm, Benedict Carling, James Marshall, Juline Savigny, Marko Storch, and Rodrigo Ledesma-Amaro. Phenotypic heterogeneity follows a growth-viability tradeoff in response to amino acid identity. Nature Communications, 15(1):6515, 2024
work page 2024
-
[4]
An experimental test of evolutionary trade-offs during temperature adaptation
Albert F Bennett and Richard E Lenski. An experimental test of evolutionary trade-offs during temperature adaptation. Proceedings of the National Academy of Sciences , 104(suppl 1):8649– 8654, 2007
work page 2007
-
[5]
Angus Buckling, Michael A Brockhurst, Michael Travisano, and Paul B Rainey. Experimental adaptation to high and low quality environments under different scales of temporal variation. Journal of Evolutionary Biology , 20(1):296–300, 2007
work page 2007
-
[6]
Maryl Lambros, Ximo Pechuan-Jorge, Daniel Biro, Kenny Ye, and Aviv Bergman. Emerging adap- tive strategies under temperature fluctuations in a laboratory evolution experiment of escherichia coli. Frontiers in microbiology, 12:724982, 2021. 12
work page 2021
-
[7]
Tuning gene expression to changing en- vironments: from rapid responses to evolutionary adaptation
Luis L´ opez-Maury, Samuel Marguerat, and J¨ urg B¨ ahler. Tuning gene expression to changing en- vironments: from rapid responses to evolutionary adaptation. Nature Reviews Genetics, 9(8):583– 593, 2008
work page 2008
-
[8]
A model for the interplay between plastic tradeoffs and evolution in changing environments
Mikhail Tikhonov, Shamit Kachru, and Daniel S Fisher. A model for the interplay between plastic tradeoffs and evolution in changing environments. Proceedings of the National Academy of Sciences, 117(16):8934–8940, 2020
work page 2020
-
[9]
Con- vergent cellular adaptation to freeze-thaw stress via a quiescence-like state in yeast
Charuhansini Tvishamayi, Farhan Ali, Nandita Chaturvedi, Nithila Madhu-Kumar, Zeenat Rashida, Chandan Muni Reddy, Ankita Ray, Stephan Herminghaus, and Shashi Thutupalli. Con- vergent cellular adaptation to freeze-thaw stress via a quiescence-like state in yeast. eLife, July 2025
work page 2025
-
[10]
Structural comparison between sucrose and trehalose in aqueous solution
Christoffer Olsson and Jan Swenson. Structural comparison between sucrose and trehalose in aqueous solution. The Journal of Physical Chemistry B , 124(15):3074–3082, 2020
work page 2020
-
[11]
Trehalose accumulation in saccharomyces cerevisiae cells: experimental data and structured modeling
Juan S Aranda, Edgar Salgado, and Patricia Taillandier. Trehalose accumulation in saccharomyces cerevisiae cells: experimental data and structured modeling. Biochemical Engineering Journal , 17(2):129–140, 2004
work page 2004
-
[12]
Hugo Tapia, Lindsey Young, Douglas Fox, Carolyn R Bertozzi, and Douglas Koshland. Increasing intracellular trehalose is sufficient to confer desiccation tolerance to saccharomyces cerevisiae. Proceedings of the National Academy of Sciences , 112(19):6122–6127, 2015
work page 2015
-
[13]
Physiological roles of trehalose in bacteria and yeasts: a comparative analysis
Juan Carlos Arg¨ uelles. Physiological roles of trehalose in bacteria and yeasts: a comparative analysis. Archives of microbiology, 174:217–224, 2000
work page 2000
-
[14]
Function of trehalose in baker’s yeast (saccharomyces cerevisiae)
Anita Panek. Function of trehalose in baker’s yeast (saccharomyces cerevisiae). Archives of Biochemistry and Biophysics , 100(3):422–425, 1963
work page 1963
-
[15]
Preser- vation of frozen yeast cells by trehalose
L Diniz-Mendes, E Bernardes, Pedro Soares De Araujo, AD Panek, and VMF Paschoalin. Preser- vation of frozen yeast cells by trehalose. Biotechnology and Bioengineering, 65(5):572–578, 1999
work page 1999
-
[16]
Lei Shi, Benjamin M Sutter, Xinyue Ye, and Benjamin P Tu. Trehalose is a key determinant of the quiescent metabolic state that fuels cell cycle progression upon return to growth. Molecular biology of the cell , 21(12):1982–1990, 2010
work page 1982
-
[17]
Monika Opalek, Bogna Smug, Michael Doebeli, and Dominika Wloch-Salamon. On the ecological significance of phenotypic heterogeneity in microbial populations undergoing starvation. Microbi- ology Spectrum, 10(1):e00450–21, 2022
work page 2022
-
[18]
The lag-phase during diauxic growth is a trade-off between fast adaptation and high growth rate
Dominique Chu and David J Barnes. The lag-phase during diauxic growth is a trade-off between fast adaptation and high growth rate. Scientific reports, 6(1):25191, 2016
work page 2016
-
[19]
A universal trade-off between growth and lag in fluctuating environments
Markus Basan, Tomoya Honda, Dimitris Christodoulou, Manuel H¨ orl, Yu-Fang Chang, Emanuele Leoncini, Avik Mukherjee, Hiroyuki Okano, Brian R Taylor, Josh M Silverman, et al. A universal trade-off between growth and lag in fluctuating environments. Nature, 584(7821):470–474, 2020
work page 2020
-
[20]
Genome-wide tran- scriptional plasticity underlies cellular adaptation to novel challenge
Shay Stern, Tali Dror, Elad Stolovicki, Naama Brenner, and Erez Braun. Genome-wide tran- scriptional plasticity underlies cellular adaptation to novel challenge. Molecular Systems Biology, 3(1):106, 2007
work page 2007
-
[21]
Nitnipa Soontorngun. Reprogramming of nonfermentative metabolism by stress-responsive tran- scription factors in the yeast saccharomyces cerevisiae. Current genetics, 63:1–7, 2017
work page 2017
-
[22]
The essence of yeast quiescence
Claudio De Virgilio. The essence of yeast quiescence. FEMS microbiology reviews, 36(2):306–339, 2012
work page 2012
-
[23]
The cell biology of quiescent yeast–a diversity of individual scenarios
Isabelle Sagot and Damien Laporte. The cell biology of quiescent yeast–a diversity of individual scenarios. Journal of Cell Science , 132(1):jcs213025, 2019
work page 2019
-
[24]
Quiescence in saccharomyces cerevisiae
Linda L Breeden and Toshio Tsukiyama. Quiescence in saccharomyces cerevisiae. Annual Review of Genetics, 56(1):253–278, 2022. 13
work page 2022
-
[25]
Anthony D Aragon, Angelina L Rodriguez, Osorio Meirelles, Sushmita Roy, George S Davidson, Phillip H Tapia, Chris Allen, Ray Joe, Don Benn, and Margaret Werner-Washburne. Charac- terization of differentiated quiescent and nonquiescent cells in yeast stationary-phase cultures. Molecular biology of the cell , 19(3):1271–1280, 2008
work page 2008
-
[26]
Proliferation/quiescence: the controversial” aller- retour”
Bertrand Daignan-Fornier and Isabelle Sagot. Proliferation/quiescence: the controversial” aller- retour”. Cell Division , 6:1–4, 2011
work page 2011
-
[27]
Staying alive: metabolic adaptations to quiescence
James R Valcourt, Johanna MS Lemons, Erin M Haley, Mina Kojima, Olukunle O Demuren, and Hilary A Coller. Staying alive: metabolic adaptations to quiescence. Cell cycle , 11(9):1680–1696, 2012
work page 2012
-
[28]
Paul K Herman. Stationary phase in yeast. Current opinion in microbiology, 5(6):602–607, 2002
work page 2002
-
[29]
Chris Allen, Sabrina B¨ uttner, Anthony D Aragon, Jason A Thomas, Osorio Meirelles, Jason E Jaetao, Don Benn, Stephanie W Ruby, Marten Veenhuis, Frank Madeo, et al. Isolation of quiescent and nonquiescent cells from yeast stationary-phase cultures.The Journal of cell biology, 174(1):89– 100, 2006
work page 2006
-
[30]
Sta- tionary phase in the yeast saccharomyces cerevisiae
Margaret Werner-Washburne, Edward Braun, Gerald C Johnston, and Richard A Singer. Sta- tionary phase in the yeast saccharomyces cerevisiae. Microbiological reviews, 57(2):383–401, 1993
work page 1993
-
[31]
Quiescence: a mechanism for escaping the effects of drug on cell populations
Tom´ as Alarc´ on and Henrik Jeldtoft Jensen. Quiescence: a mechanism for escaping the effects of drug on cell populations. Journal of The Royal Society Interface , 8(54):99–106, 2011
work page 2011
-
[32]
Exit from quiescence displays a memory of cell growth and division
Xia Wang, Kotaro Fujimaki, Geoffrey C Mitchell, Jungeun Sarah Kwon, Kimiko Della Croce, Chris Langsdorf, Hao Helen Zhang, and Guang Yao. Exit from quiescence displays a memory of cell growth and division. Nature communications, 8(1):321, 2017
work page 2017
-
[33]
Cellular quiescence in budding yeast
Siyu Sun and David Gresham. Cellular quiescence in budding yeast. Yeast, 38(1):12–29, 2021
work page 2021
-
[34]
Observation of universal ageing dynamics in an- tibiotic persistence
Yoav Kaplan, Shaked Reich, Elyaqim Oster, Shani Maoz, Irit Levin-Reisman, Irine Ronin, Orit Gefen, Oded Agam, and Nathalie Q Balaban. Observation of universal ageing dynamics in an- tibiotic persistence. Nature, 600(7888):290–294, 2021
work page 2021
-
[35]
Gabrielle Woronoff, Philippe Nghe, Jean Baudry, Laurent Boitard, Erez Braun, Andrew D Grif- fiths, and J´ erˆ ome Bibette. Metabolic cost of rapid adaptation of single yeast cells.Proceedings of the National Academy of Sciences , 117(20):10660–10666, 2020
work page 2020
-
[36]
Shaping of microbial phenotypes by trade-offs
Manlu Zhu and Xiongfeng Dai. Shaping of microbial phenotypes by trade-offs. Nature Commu- nications, 15(1):4238, 2024
work page 2024
-
[37]
Frank J Bruggeman, Bas Teusink, and Ralf Steuer. Trade-offs between the instantaneous growth rate and long-term fitness: consequences for microbial physiology and predictive computational models. Bioessays, 45(10):2300015, 2023
work page 2023
-
[38]
Leonie J Jahn, Christian Munck, Mostafa MH Ellabaan, and Morten OA Sommer. Adaptive labo- ratory evolution of antibiotic resistance using different selection regimes lead to similar phenotypes and genotypes. Frontiers in microbiology, 8:816, 2017
work page 2017
-
[39]
Selection of resistance at lethal and non-lethal antibiotic concentrations
Diarmaid Hughes and Dan I Andersson. Selection of resistance at lethal and non-lethal antibiotic concentrations. Current opinion in microbiology , 15(5):555–560, 2012
work page 2012
-
[40]
Selection of antibiotic resistance at very low antibiotic concentrations
Linus Sandegren. Selection of antibiotic resistance at very low antibiotic concentrations. Upsala journal of medical sciences , 119(2):103–107, 2014
work page 2014
-
[41]
Trade-offs between microbial growth phases lead to frequency-dependent and non-transitive selection
Michael Manhart, Bharat V Adkar, and Eugene I Shakhnovich. Trade-offs between microbial growth phases lead to frequency-dependent and non-transitive selection. Proceedings of the Royal Society B: Biological Sciences, 285(1872):20172459, 2018
work page 2018
-
[42]
Wide lag time distributions break a trade-off between repro- duction and survival in bacteria
Stefany Moreno-G´ amez, Daniel J Kiviet, Cl´ ement Vulin, Susan Schlegel, Kim Schlegel, G Sander van Doorn, and Martin Ackermann. Wide lag time distributions break a trade-off between repro- duction and survival in bacteria. Proceedings of the National Academy of Sciences, 117(31):18729– 18736, 2020. 14
work page 2020
-
[43]
Neutral drift upon threshold-like selection promotes variation in antibiotic resistance phenotype
Ay¸ se Nisan Erdo˘ gan, Pouria Dasmeh, Raymond D Socha, John Z Chen, Benjamin E Life, Rachel Jun, Linda Kiritchkov, Dan Kehila, Adrian WR Serohijos, and Nobuhiko Tokuriki. Neutral drift upon threshold-like selection promotes variation in antibiotic resistance phenotype. Nature Communications, 15(1):10813, 2024
work page 2024
-
[44]
Shimon Bershtein, Adrian WR Serohijos, and Eugene I Shakhnovich. Bridging the physical scales in evolutionary biology: from protein sequence space to fitness of organisms and populations. Current opinion in structural biology , 42:31–40, 2017
work page 2017
-
[45]
Levels of selection on threshold characters
Jacob A Moorad and Timothy A Linksvayer. Levels of selection on threshold characters. Genetics, 179(2):899–905, 2008
work page 2008
-
[46]
Estimating heritability in a threshold trait: heat-shock tolerance in drosophila buzzatii
Robert A Krebs and Volker Loeschcke. Estimating heritability in a threshold trait: heat-shock tolerance in drosophila buzzatii. Heredity, 79(3):252–259, 1997
work page 1997
-
[47]
Multiple means to the same end: the genetic basis of acquired stress resistance in yeast
David B Berry, Qiaoning Guan, James Hose, Suraiya Haroon, Marinella Gebbia, Lawrence E Heisler, Corey Nislow, Guri Giaever, and Audrey P Gasch. Multiple means to the same end: the genetic basis of acquired stress resistance in yeast. PLoS genetics, 7(11):e1002353, 2011
work page 2011
-
[48]
Stress-induced variation in evolution: from behavioural plasticity to genetic assimilation
Alexander V Badyaev. Stress-induced variation in evolution: from behavioural plasticity to genetic assimilation. Proceedings of the Royal Society B: Biological Sciences , 272(1566):877–886, 2005
work page 2005
-
[49]
Phenotypic plasticity and evolution by genetic assimilation
Vidyanand Nanjundiah. Phenotypic plasticity and evolution by genetic assimilation. Origination of organismal form: Beyond the gene in developmental and evolutionary biology , 2:245, 2003
work page 2003
-
[50]
Bacterial persister cell formation and dormancy
Thomas K Wood, Stephen J Knabel, and Brian W Kwan. Bacterial persister cell formation and dormancy. Applied and environmental microbiology, 79(23):7116–7121, 2013
work page 2013
-
[51]
Bacterial persistence: a model of survival in changing environments
Edo Kussell, Roy Kishony, Nathalie Q Balaban, and Stanislas Leibler. Bacterial persistence: a model of survival in changing environments. Genetics, 169(4):1807–1814, 2005
work page 2005
-
[52]
Bacterial persistence as a phenotypic switch
Nathalie Q Balaban, Jack Merrin, Remy Chait, Lukasz Kowalik, and Stanislas Leibler. Bacterial persistence as a phenotypic switch. Science, 305(5690):1622–1625, 2004
work page 2004
-
[53]
Bram Van den Bergh, Joran E Michiels, Tom Wenseleers, Etthel M Windels, Pieterjan Vanden Boer, Donaat Kestemont, Luc De Meester, Kevin J Verstrepen, Natalie Verstraeten, Maarten Fau- vart, et al. Frequency of antibiotic application drives rapid evolutionary adaptation of escherichia coli persistence. Nature microbiology, 1(5):1–7, 2016
work page 2016
-
[54]
Optimization of lag time underlies antibiotic tolerance in evolved bacterial populations
Ofer Fridman, Amir Goldberg, Irine Ronin, Noam Shoresh, and Nathalie Q Balaban. Optimization of lag time underlies antibiotic tolerance in evolved bacterial populations. Nature, 513(7518):418– 421, 2014
work page 2014
-
[55]
A systematic review on quiescent state research approaches in s
Monika Opalek, Hanna Tutaj, Adrian Pirog, Bogna J Smug, Joanna Rutkowska, and Dominika Wloch-Salamon. A systematic review on quiescent state research approaches in s. cerevisiae.Cells, 12(12):1608, 2023
work page 2023
-
[56]
Gregory J Velicer and Richard E Lenski. Evolutionary trade-offs under conditions of resource abundance and scarcity: experiments with bacteria. Ecology, 80(4):1168–1179, 1999
work page 1999
-
[57]
Minna Pekkonen, Tarmo Ketola, and Jouni T Laakso. Resource availability and competition shape the evolution of survival and growth ability in a bacterial community. PLoS One, 8(9):e76471, 2013
work page 2013
-
[58]
Dynamics of bacterial populations under the feast-famine cycles
Yusuke Himeoka and Namiko Mitarai. Dynamics of bacterial populations under the feast-famine cycles. Physical Review Research, 2(1):013372, 2020
work page 2020
-
[59]
Population dynamics of general- ist/specialist strategies in the feast-famine cycle
Rintaro Niimi, Chikara Furusawa, and Yusuke Himeoka. Population dynamics of general- ist/specialist strategies in the feast-famine cycle. bioRxiv, pages 2025–05, 2025
work page 2025
-
[60]
Silja Borring L˚ astad and Namiko Mitarai. Triggered and spontaneous dormancy in bacteria during feast-famine cycles with stochastic antibiotic application. bioRxiv, pages 2025–02, 2025. 15
work page 2025
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.