Motor shot noise explains active fluctuations in a single cilium
Pith reviewed 2026-05-16 23:23 UTC · model grok-4.3
The pith
Fluctuations in the number of bound motors fully explain the observed variability in a single cilium's beat cycle.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Fluctuations in the number of bound motors are sufficient to explain experimentally observed fluctuations in the cilia beat, including a quality factor Q that measures oscillation precision and phase defects of intra-cilium synchronization.
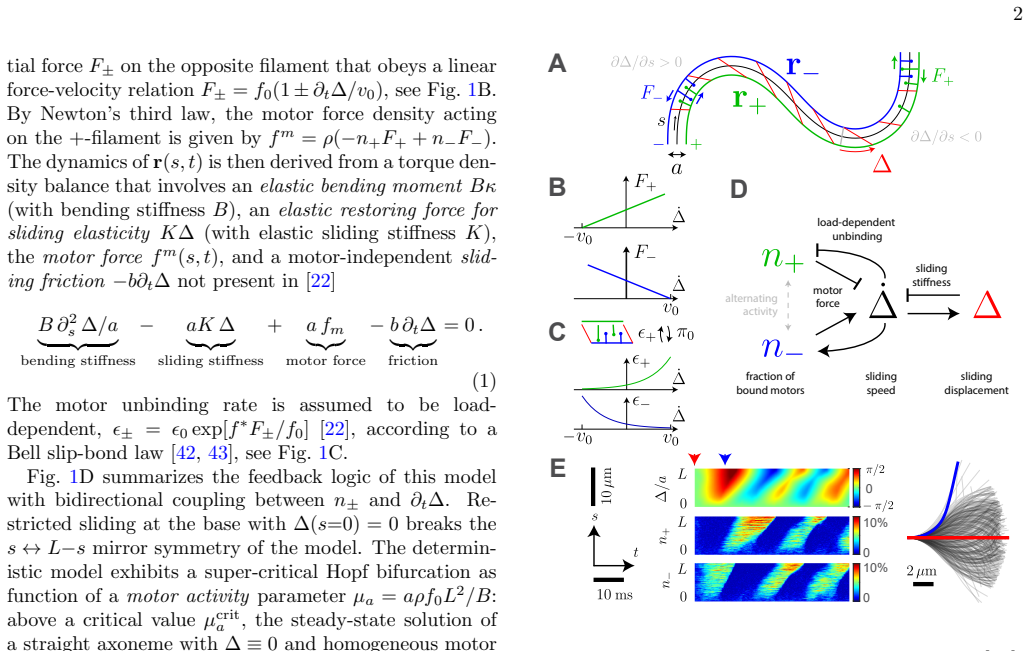
What carries the argument
Motor shot noise from stochastic binding and unbinding of a small number of dynein motors, mapped through a mechanical model of the axoneme to the resulting beat dynamics.
If this is right
- Theories of motor control must incorporate intrinsic shot noise rather than assuming additional regulatory feedback to achieve observed precision.
- The direct quantitative link between microscopic motor statistics and mesoscopic beat fluctuations is established without intermediate mechanisms.
- Similar motor-number fluctuations are expected to limit regularity in other active biological oscillators driven by small numbers of motors.
- Experimental variation of motor density should produce predictable changes in the quality factor of the beat.
Where Pith is reading between the lines
- If the mapping holds, altering motor expression levels in live cells should produce measurable shifts in beat regularity that can be checked against the model's predictions.
- The same counting-noise mechanism may set baseline variability in related systems such as flagellar propulsion or cytoskeletal networks.
- Models of coordinated beating across multiple cilia in tissues would need to treat single-cilium motor shot noise as an irreducible source of phase jitter.
Load-bearing premise
Fluctuations in the number of bound motors dominate over all other sources of noise and the mapping from motor statistics to beat mechanics captures real ciliary dynamics without extra fitted mechanisms.
What would settle it
Direct measurement of the number of bound motors during beating combined with a mismatch between the predicted quality factor from those counts and the experimentally observed Q would falsify the claim.
Figures


read the original abstract
Mesoscopic fluctuations reveal stochastic dynamics of molecules in both inanimate and living matter. We investigate how small-number fluctuations shape the collective dynamics of molecular motors using motile cilia as model system. We theoretically show that fluctuations in the number of bound motors are sufficient to explain experimentally observed fluctuations in the cilia beat, including a quality factor $Q$ that measures oscillation precision and phase defects of intra-cilium synchronization. Our findings constrain theories of motor control and establish a link between microscopic motor noise and mesoscopic non-equilibrium dynamics.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that fluctuations in the number of bound motors (motor shot noise) are sufficient to explain experimentally observed fluctuations in the beat of a single cilium. This includes accounting for the quality factor Q measuring oscillation precision and the statistics of phase defects arising from intra-cilium synchronization. The work uses a reduced-order model in which Poisson statistics of bound-motor number N(t) enter an effective driving force or curvature term, thereby linking microscopic motor noise to mesoscopic non-equilibrium dynamics without additional control mechanisms.
Significance. If the result holds, the paper would establish a direct, parameter-light connection between small-number motor stochasticity and the precision of ciliary beating, constraining theories of motor regulation within the axoneme. It offers a parsimonious explanation for active fluctuations and highlights the role of shot noise in biological oscillators, with potential implications for other systems involving collective motor dynamics.
major comments (2)
- [Theoretical model (derivation of effective oscillator)] The sufficiency claim requires demonstrating that variance in bound-motor number N(t) dominates other noise channels (ATP binding kinetics, stepping noise, elastic compliance, hydrodynamic drag). No explicit variance decomposition or sensitivity analysis is provided to quantify their relative sizes, leaving the load-bearing assumption untested.
- [Results and comparison to experiment] No control is presented in which N is held fixed while retaining stochastic binding or stepping rates, to verify whether the measured Q and phase-defect statistics remain unchanged. This test is necessary to isolate the motor-number contribution and support the assertion of sufficiency.
minor comments (1)
- [Model equations] Clarify the precise functional form of the mapping from N(t) to the curvature or driving term in the reduced-order model, including any assumptions about linearity or weak nonlinearity at observed amplitudes.
Simulated Author's Rebuttal
We thank the referee for their careful reading and constructive comments on our manuscript. We address each major comment below and outline the revisions we will make to strengthen the presentation.
read point-by-point responses
-
Referee: [Theoretical model (derivation of effective oscillator)] The sufficiency claim requires demonstrating that variance in bound-motor number N(t) dominates other noise channels (ATP binding kinetics, stepping noise, elastic compliance, hydrodynamic drag). No explicit variance decomposition or sensitivity analysis is provided to quantify their relative sizes, leaving the load-bearing assumption untested.
Authors: We thank the referee for this observation. Our reduced-order model intentionally coarse-grains microscopic processes (including ATP kinetics and stepping) into an effective curvature drive whose fluctuations are driven by Poisson statistics of N(t). This isolates the contribution of motor-number shot noise without claiming it is the sole noise source in the axoneme. To address the concern, the revised manuscript will include a brief sensitivity analysis in which we vary the effective parameters associated with other noise channels (e.g., drag coefficient and elastic stiffness) while keeping the variance of N fixed; the results show that Q and phase-defect statistics remain largely unchanged, supporting the dominance of motor-number fluctuations within the model’s scope. revision: partial
-
Referee: [Results and comparison to experiment] No control is presented in which N is held fixed while retaining stochastic binding or stepping rates, to verify whether the measured Q and phase-defect statistics remain unchanged. This test is necessary to isolate the motor-number contribution and support the assertion of sufficiency.
Authors: We agree that an explicit control would help isolate the role of number fluctuations. In the revised manuscript we will add a mean-field comparison in which N is replaced by its deterministic average while the underlying binding kinetics remain stochastic; the resulting deterministic drive produces a significantly higher Q and eliminates the observed phase defects. This control confirms that the fluctuations in Q and synchronization arise specifically from the stochastic variation in N(t) rather than from other stochastic rates alone. revision: yes
Circularity Check
No significant circularity; derivation uses independent motor-kinetics model to predict Q
full rationale
The paper models bound-motor number N(t) as a Poisson process with independent rates for attachment/detachment, then inserts the resulting force fluctuations into a reduced-order oscillator equation for the cilium beat. The quality factor Q and phase jitter emerge as derived statistics from the variance of this driving term under stated assumptions about axonemal stiffness and hydrodynamics. No equation reduces to a fitted parameter renamed as prediction, no self-citation chain supplies the central mapping, and the abstract's claim of sufficiency is presented as a forward calculation rather than a tautology. The derivation chain is therefore self-contained against external motor-kinetic parameters.
Axiom & Free-Parameter Ledger
free parameters (1)
- effective number of motors or binding rate
axioms (1)
- domain assumption Motor attachment and detachment follow Poisson statistics (shot noise).
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
fluctuations in the number of bound motors are sufficient to explain experimentally observed fluctuations in the cilia beat, including a quality factor Q ... phase defects of intra-cilium synchronization
-
IndisputableMonolith/Foundation/AlexanderDuality.leanalexander_duality_circle_linking unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
N cilium ≈1.7·10^4 ... Q scales asymptotically as Q∼N
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Kubo, The fluctuation-dissipation theorem, Reports on Progress in Physics29, 255 (1966)
R. Kubo, The fluctuation-dissipation theorem, Reports on Progress in Physics29, 255 (1966)
work page 1966
-
[2]
C. P. Brangwynne, G. H. Koenderink, F. C. MacKin- tosh, and D. A. Weitz, Nonequilibrium microtubule fluc- tuations in a model cytoskeleton, Phys. Rev. Lett.100, 118104 (2008)
work page 2008
- [3]
-
[4]
F.-Y. Chu, S. C. Haley, and A. Zidovska, On the origin of shape fluctuations of the cell nucleus, Proc. Natl. Acad. Sci. U.S.A.114, 10338 (2017)
work page 2017
-
[5]
H. Turlier, D. A. Fedosov, B. Audoly, T. Auth, N. S. Gov, C. Sykes, J.-F. Joanny, G. Gompper, and T. Betz, Equilibrium physics breakdown reveals the active nature of red blood cell flickering, Nature physics12, 513 (2016)
work page 2016
-
[6]
H. C. Berg, The rotary motor of bacterial flagella, Annual review of biochemistry72, 19 (2003)
work page 2003
-
[7]
A. Samuel and H. C. Berg, Fluctuation analysis of rota- tional speeds of the bacterial flagellar motor., Proc. Natl. Acad. Sci. U.S.A.92, 3502 (1995)
work page 1995
-
[8]
A. F. Huxley and R. Niedergerke, Structural changes in muscle during contraction: interference microscopy of liv- ing muscle fibres, Nature173, 971 (1954)
work page 1954
-
[9]
D. Haertter, L. Hauke, T. Driehorst, K. Nishi, W.- H. Zimmermann, and C. F. Schmidt, Stochastic tug-of- war among sarcomeres mediates cardiomyocyte response to environmental stiffness, eLife 10.7554/elife.97321.1 (2024)
-
[10]
A. J. Hudspeth, Mechanical amplification of stimuli by hair cells, Current opinion in neurobiology7, 480 (1997)
work page 1997
-
[11]
B. Nadrowski, P. Martin, and F. J¨ ulicher, Active hair- bundle motility harnesses noise to operate near an opti- mum of mechanosensitivity, Proc. Natl. Acad. Sci. U.S.A. 101, 12195 (2004)
work page 2004
-
[12]
Gray,Ciliary movement(Cambridge University Press, 1928)
J. Gray,Ciliary movement(Cambridge University Press, 1928)
work page 1928
-
[13]
M. A. Sleigh,The Biology of Cilia and Flagella(Perga- mon, Oxford, 1962)
work page 1962
-
[14]
D. R. Mitchell, Evolution of cilia, Cold Spring Harbor Perspectives in Biology9, a028290 (2017)
work page 2017
- [15]
-
[16]
C. B. Lindemann, Testing the geometric clutch hypoth- esis, Biology of the Cell96, 681 (2004)
work page 2004
-
[17]
C. J. Brokaw, Thinking about flagellar oscillation, Cell Motility and the Cytoskeleton66, 425 (2009)
work page 2009
-
[18]
S. Camalet and F. J¨ ulicher, Generic aspects of axonemal beating, New Journal of Physics2, 324 (2000)
work page 2000
-
[19]
I. H. Riedel-Kruse, A. Hilfinger, J. Howard, and F. J¨ ulicher, How molecular motors shape the flagellar beat, HFSP journal1, 192 (2007)
work page 2007
- [20]
-
[21]
V. F. Geyer, J. Howard, and P. Sartori, Ciliary beating patterns map onto a low-dimensional behavioural space, Nature Physics 2022 18:318, 332 (2022)
work page 2022
-
[22]
J. F. Cass and H. Bloomfield-Gadˆ elha, The reaction- diffusion basis of animated patterns in eukaryotic flagella, Nature Communications14, 5638 (2023)
work page 2023
-
[23]
P. Sartori, V. F. Geyer, A. Scholich, F. J¨ ulicher, and J. Howard, Dynamic curvature regulation accounts for the symmetric and asymmetric beats ofChlamydomonas flagella, eLife5, e13258 (2016)
work page 2016
-
[24]
P. Bayly and S. Dutcher, Steady dynein forces induce flutter instability and propagating waves in mathematical models of flagella, Journal of The Royal Society Interface 13, 20160523 (2016)
work page 2016
-
[25]
B. Chakrabarti and D. Saintillan, Spontaneous oscilla- tions, beating patterns, and hydrodynamics of active mi- crofilaments, Physical Review Fluids4, 043102 (2019)
work page 2019
- [26]
- [27]
-
[28]
R. E. Goldstein, M. Polin, and I. Tuval, Noise and syn- chronization in pairs of beating eukaryotic flagella, Phys. Rev. Lett.103, 168103 (2009). 6
work page 2009
-
[29]
R. Ma, G. S. Klindt, I. H. Riedel-Kruse, F. J¨ ulicher, and B. M. Friedrich, Active phase and amplitude fluctuations of flagellar beating, Phys. Rev. Lett.113, 048101 (2014)
work page 2014
-
[30]
K. Y. Wan and R. E. Goldstein, Rhythmicity, recurrence, and recovery of flagellar beating, Phys. Rev. Lett.113, 238103 (2014)
work page 2014
- [31]
-
[32]
R. E. Goldstein, M. Polin, and I. Tuval, Emergence of synchronized beating during the regrowth of eukaryotic flagella, Phys. Rev. Lett.107, 148103 (2011)
work page 2011
-
[33]
A. Solovev and B. M. Friedrich, Synchronization in cilia carpets and the Kuramoto model with local coupling: Breakup of global synchronization in the presence of noise, Chaos: An Interdisciplinary Journal of Nonlinear Science32(2022)
work page 2022
-
[34]
G. Costantini and A. Puglisi, Thermodynamic precision of a chain of motors: the difference between phase and noise correlation, Journal of Statistical Mechanics: The- ory and Experiment2024, 024003 (2024)
work page 2024
- [35]
-
[36]
I. S. Aranson and L. Kramer, The world of the com- plex ginzburg-landau equation, Rev. Mod. Phys.74, 99 (2002), publisher: American Physical Society
work page 2002
-
[37]
P.-Y. Pla¸ cais, M. Balland, T. Gu´ erin, J.-F. Joanny, and P. Martin, Spontaneous oscillations of a minimal acto- myosin system under elastic loading, Phys. Rev. Lett. 103, 158102 (2009)
work page 2009
-
[38]
T. Gu´ erin, J. Prost, and J.-F. Joanny, Dynamical behav- ior of molecular motor assemblies in the rigid and cross- bridge models, The European Physical Journal E34, 60 (2011)
work page 2011
-
[39]
G. S. Klindt, C. Ruloff, C. Wagner, and B. M. Friedrich, Load response of the flagellar beat, Phys. Rev. Lett.117, 258101 (2016)
work page 2016
-
[40]
N. Pellicciotta, E. Hamilton, J. Kotar, M. Faucourt, N. Delgehyr, N. Spassky, and P. Cicuta, Entrainment of mammalian motile cilia in the brain with hydrodynamic forces, Proc. Natl. Acad. Sci. U.S.A.117, 8315 (2020)
work page 2020
- [41]
-
[42]
G. I. Bell, Models for the specific adhesion of cells to cells, Science200, 618 (1978)
work page 1978
-
[43]
H. A. Kramers, Brownian motion in a field of force and the diffusion model of chemical reactions, Physica7, 284 (1940)
work page 1940
-
[44]
B. Rallabandi, Q. Wang, and M. Potomkin, Self- sustained three-dimensional beating of a model eukary- otic flagellum, Soft Matter18, 5312 (2022)
work page 2022
-
[45]
J. Howard and R. Clark, Mechanics of motor proteins and the cytoskeleton, Appl. Mech. Rev.55, B39 (2002)
work page 2002
-
[46]
A. C. Barato and U. Seifert, Thermodynamic uncertainty relation for biomolecular processes, Phys. Rev. Lett.114, 158101 (2015)
work page 2015
-
[47]
G. S. Klindt and B. M. Friedrich, Flagellar swimmers os- cillate between pusher-and puller-type swimming, Phys. Rev. E92, 063019 (2015)
work page 2015
-
[48]
B. M. Friedrich, Load response of shape-changing mi- croswimmers scales with their swimming efficiency, Phys. Rev. E97, 042416 (2018)
work page 2018
-
[49]
U. R¨ uffer and W. Nultsch, Flagellar photoresponses of Chlamydomonascells held on micropipettes: III. Shock response, Botanica Acta108, 255 (1995)
work page 1995
-
[50]
K. Y. Wan and R. E. Goldstein, Time irreversibility and criticality in the motility of a flagellate microorganism, Phys. Rev. Lett.121, 058103 (2018)
work page 2018
-
[51]
M. R. Miller, N. Mannowetz, A. T. Iavarone, R. Safavi, E. O. Gracheva, J. F. Smith, R. Z. Hill, D. M. Bautista, Y. Kirichok, and P. V. Lishko, Unconventional endo- cannabinoid signaling governs sperm activation via the sex hormone progesterone, Science352, 555 (2016)
work page 2016
-
[52]
S. Veeraragavan, F. Y. Parast, R. Nosrati, and R. Prab- hakar, Elastohydrodynamic mechanisms govern beat pat- tern transitions in eukaryotic flagella, bioRxiv , 2024 (2024)
work page 2024
-
[53]
Kuramoto,Chemical Oscillations, Waves, and Turbu- lence(Springer-Verlag, Berlin, Heidelberg, 1984)
Y. Kuramoto,Chemical Oscillations, Waves, and Turbu- lence(Springer-Verlag, Berlin, Heidelberg, 1984)
work page 1984
-
[54]
I. H. Riedel-Kruse, H. Andreas, H. Jonathon, and F. J¨ ulicher, How molecular motors shape the flagellar beat, HFSP Journal1, 192 (2007)
work page 2007
-
[55]
P. Sartori, V. F. Geyer, A. Scholich, F. J¨ ulicher, and J. Howard, Dynamic curvature regulation accounts for the symmetric and asymmetric beats of chlamydomonas flagella, eLife5, e13258 (2016), publisher: eLife Sciences Publications, Ltd
work page 2016
-
[56]
Howard, Mechanical signaling in networks of motor and cytoskeletal proteins, Ann
J. Howard, Mechanical signaling in networks of motor and cytoskeletal proteins, Ann. Rev. Biophys.38, 217 (2009)
work page 2009
-
[57]
B. Kralemann, L. Cimponeriu, M. Rosenblum, A. Pikovsky, and R. Mrowka, Phase dynamics of coupled oscillators reconstructed from data, Phys. Rev. E77, 066205 (2008)
work page 2008
-
[58]
J. F. Cass and H. Bloomfield-Gadˆ elha, Predicting mi- croscale beat patterns from nanoscale chemomechanics in eukaryotic flagella, bioRxiv: 2024.08.14.607876 , 2024 (2024)
work page 2024
-
[59]
G. Xu, K. S. Wilson, R. J. Okamoto, J.-Y. Shao, S. K. Dutcher, and P. V. Bayly, Flexural rigidity and shear stiffness of flagella estimated from induced bends and counterbends, Biophysical journal110, 2759 (2016)
work page 2016
-
[60]
M. Okuno and Y. Hiramoto, Direct measurements of the stiffness of echinoderm sperm flagella, Journal of Exper- imental Biology79, 235 (1979)
work page 1979
-
[61]
M. Okuno, Inhibition and relaxation of sea urchin sperm flagella by vanadate, Journal of Cell Biology85, 712 (1980)
work page 1980
-
[62]
E. H. Lee, X. Ouyang, and J. Howard, The wavelength of the ciliary beat in chlamydomonas saturates at long ciliary lengths, Biophysical Journal124, 2961 (2025)
work page 2025
-
[63]
J. Gray and G. Hancock, The propulsion of sea-urchin spermatozoa, Journal of Experimental Biology32, 802 (1955)
work page 1955
-
[64]
B. M. Friedrich, I. H. Riedel-Kruse, J. Howard, and F. J¨ ulicher, High-precision tracking of sperm swimming fine structure provides strong test of resistive force the- ory, Journal of Experimental Biology213, 1226 (2010)
work page 2010
-
[65]
J. Happel and H. Brenner,Low Reynolds number hydro- dynamics: with special applications to particulate media, Vol. 1 (Springer Science & Business Media, 2012)
work page 2012
-
[66]
V. F. Geyer, P. Sartori, B. M. Friedrich, F. J¨ ulicher, and J. Howard, Independent control of the static and dynamic components of theChlamydomonasflagellar beat, Cur- rent Biology26, 1098 (2016). 7 END MATTER Non-dimensional parameters.The stochastic model depends on 7 non-dimensional parameters µa =aρf 0L2/B , µ=a 2KL2/B , η=π 0τ , ζ=a/(v 0τ), f ∗ =f 0...
-
[67]
Movie captions for Supplemental Movies M1 to M4
-
[68]
Supplemental Data Analysis Methods
-
[69]
Supplemental Information on Mathematical Model and Numerical Methods Supplemental Movie M1.Experimental data of beating axoneme without phase defect from experiment without motor extraction [15].Leftmost:kymograph of local phaseφ(s, t),left-center:tangent angle profileγ(s, t) at time point indicated by vertical line in first panel,right-center:tangent ang...
work page 2000
-
[70]
Therefore, in this first step, we restrict ourselves to short simulations forN remain/N= 72%
For most parameter setsθ, we observe either no oscillation at all, or standing wave oscillations withλ≫L. Therefore, in this first step, we restrict ourselves to short simulations forN remain/N= 72%. The first parameter sets were chosen from a prior distribution, chosen as a multi-variate Gaussian distribution centered at the parameters from Cass et al. [...
-
[71]
S5.Non-isochrony of cilia beating
In a second step, we proceed analogous to the first step, while performing slightly longer, yet still short sim- ulations, now both without and with motor extraction, forN remain/N= 100% andN remain/N= 72%, re- Kotzet al.— Supplemental Material S16 80% 90% 100% 110% 120% Relative instantaneous amplitude 80% 85% 90% 95% 100% 105% 110% 115% 120%Relative ins...
-
[72]
In the third step, we perform even longer simulations to accurately determine the quality factorsQ 72% and Q100%. In a final step, we determine the maximum-likelihood estimate for model parameters, ˆθ= argmaxl(θ), usingl(θ) = l3(θ) as determined after the third step, and computex( ˆθ) with same accuracy as used for the main numerical results reported in F...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.