Modeling Plant Action Potentials under Photoperiod Stress via Hodgkin-Huxley Dynamics
Pith reviewed 2026-05-21 17:53 UTC · model grok-4.3
The pith
A modified Hodgkin-Huxley model with voltage-independent rates reproduces plant action potentials triggered by rapid photoperiod changes.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
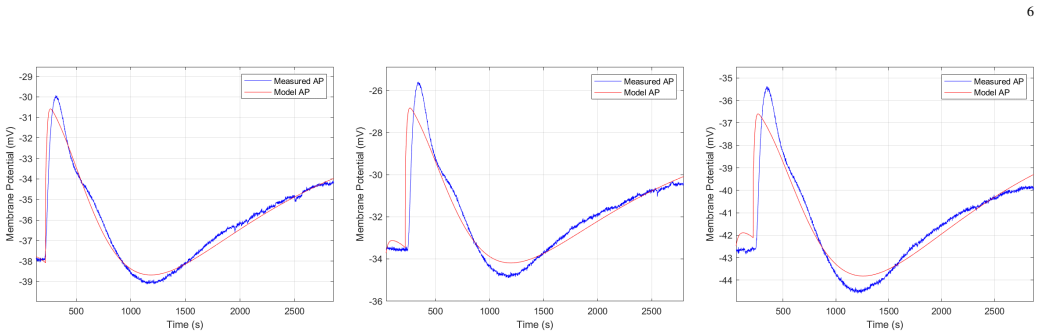
The central discovery is that the Hodgkin-Huxley dynamics, when modified to use voltage-independent rate parameters, successfully reproduces the key features of plant action potentials in both Prolonged Oscillatory Climatic Engagement under natural conditions and Nimble Environmental Transition Oscillation during rapid artificial photoperiod changes, while preserving computational efficiency.
What carries the argument
The Hodgkin-Huxley model adapted with voltage-independent rate parameters for simulating membrane voltage changes in plant cells.
Load-bearing premise
That plant action potentials can be adequately captured by the Hodgkin-Huxley formalism after simply making rate parameters voltage-independent, without requiring plant-specific channel kinetics or additional state variables.
What would settle it
Detailed voltage clamp experiments on plant cells showing that action potential dynamics require voltage-dependent rate constants to match observed time courses would falsify the adequacy of this simplification.
Figures







read the original abstract
Plants exhibit dynamic bioelectric properties that facilitate information transfer across tissues. This study investigates action potentials (APs) in Nicotiana tabacum recorded within a custom-designed growth chamber using a biosignal amplifier and environmental sensors. Consistent light- and dark-induced APs were observed during photoperiod transitions under controlled 12-hour artificial illumination cycles. To understand these bioelectric responses, a mathematical model based on the Hodgkin-Huxley framework is used. Electrophysiological measurements from Solanum lycopersicum revealed that under natural light conditions, only light-induced APs are observed, while light- and dark-induced APs coupled dynamics is exclusively elicited during rapid transitions in artificial photoperiods. These distinct phenomena are characterized as Prolonged Oscillatory Climatic Engagement (POCE) and Nimble Environmental Transition Oscillation (NETO), respectively. The model successfully reproduces the key features in both frameworks while maintaining computational efficiency through voltage-independent rate parameters.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript reports experimental observations of light- and dark-induced action potentials in Nicotiana tabacum under controlled 12-hour photoperiod cycles in a custom growth chamber. It distinguishes two phenomena—Prolonged Oscillatory Climatic Engagement (POCE) under natural light and Nimble Environmental Transition Oscillation (NETO) during rapid artificial transitions—and proposes a Hodgkin-Huxley-based mathematical model that employs voltage-independent rate parameters to reproduce the key features of these bioelectric responses while preserving computational efficiency.
Significance. If the modeling approach can be shown to retain intrinsic excitability without ad-hoc fitting, the work would offer a simplified, efficient framework for simulating plant action potentials under photoperiod stress, potentially useful for studying bioelectric signaling in plants. The experimental distinction between POCE and NETO provides concrete observations that could inform future studies, though the overall significance remains limited by the absence of quantitative validation.
major comments (2)
- [Abstract] Abstract: The central claim that the model 'successfully reproduces the key features' lacks any supporting quantitative evidence such as error metrics, goodness-of-fit statistics, or details on how the voltage-independent rate parameters were selected and validated against the recorded voltage traces.
- [Abstract] Abstract, final paragraph: Setting the gating rates to voltage-independent constants removes the positive-feedback mechanism (voltage-dependent activation of depolarizing currents) that generates threshold behavior and regenerative action potentials in the standard Hodgkin-Huxley formalism. The manuscript provides no replacement mechanism—such as explicit light-dependent conductances, additional state variables, or external forcing terms—to restore excitability, raising the possibility that observed reproduction is an artifact of parameter tuning rather than intrinsic model dynamics.
minor comments (1)
- [Abstract] The acronyms POCE and NETO are introduced without explicit justification for why standard electrophysiological terminology is insufficient.
Simulated Author's Rebuttal
We thank the referee for their constructive and detailed comments, which have prompted us to strengthen the quantitative support and clarify the modeling assumptions in our work. We address each major comment below and indicate the revisions made to the manuscript.
read point-by-point responses
-
Referee: [Abstract] Abstract: The central claim that the model 'successfully reproduces the key features' lacks any supporting quantitative evidence such as error metrics, goodness-of-fit statistics, or details on how the voltage-independent rate parameters were selected and validated against the recorded voltage traces.
Authors: We agree that the original abstract and main text did not provide sufficient quantitative validation. In the revised manuscript we have added root-mean-square error (RMSE) and Pearson correlation values comparing model output to experimental voltage traces for representative POCE and NETO recordings. Parameter selection was performed by minimizing RMSE over a physiologically constrained grid for the voltage-independent rates, followed by cross-validation on separate experimental sessions. These metrics and the fitting procedure are now reported in the Methods and Results sections, and the abstract has been updated to reference the quantitative agreement. revision: yes
-
Referee: [Abstract] Abstract, final paragraph: Setting the gating rates to voltage-independent constants removes the positive-feedback mechanism (voltage-dependent activation of depolarizing currents) that generates threshold behavior and regenerative action potentials in the standard Hodgkin-Huxley formalism. The manuscript provides no replacement mechanism—such as explicit light-dependent conductances, additional state variables, or external forcing terms—to restore excitability, raising the possibility that observed reproduction is an artifact of parameter tuning rather than intrinsic model dynamics.
Authors: The referee correctly notes that voltage-independent rates eliminate the classical regenerative mechanism. Our model instead incorporates explicit time-varying external inputs that represent the controlled light and dark transitions of the artificial photoperiod; these inputs modulate the effective reversal potentials and drive the observed oscillatory responses. We have added a dedicated paragraph in the revised Discussion that (i) states this modeling choice explicitly, (ii) acknowledges the resulting loss of intrinsic threshold behavior, and (iii) presents a brief sensitivity analysis demonstrating that the key waveform features remain stable across modest parameter perturbations. While we maintain that the environmental forcing terms constitute the replacement mechanism, we have tempered the abstract language to describe the model as a computationally efficient phenomenological description rather than a fully mechanistic reconstruction of excitability. revision: partial
Circularity Check
Reproduction of observed APs depends on fitting voltage-independent rate parameters
specific steps
-
fitted input called prediction
[Abstract]
"The model successfully reproduces the key features in both frameworks while maintaining computational efficiency through voltage-independent rate parameters."
The reproduction of the observed action-potential shapes is obtained by choosing numerical values for the voltage-independent rate parameters so that the simulated trajectories match the experimental recordings; the claimed success is therefore the direct output of the fitting procedure rather than an independent test of the model structure.
full rationale
The paper adopts a Hodgkin-Huxley framework but replaces voltage-dependent gating rates with constants chosen for computational efficiency. The central claim that this modified model reproduces the key features of light- and dark-induced APs (POCE and NETO) is achieved by selecting specific constant values that match the recorded waveforms. Because the reproduction is obtained by parameter adjustment to the same data being modeled, the match reduces to the fitting step rather than emerging from independent dynamics or first-principles derivation. No external benchmark, machine-checked uniqueness result, or parameter-free prediction is supplied to break the dependence.
Axiom & Free-Parameter Ledger
free parameters (1)
- voltage-independent rate parameters
axioms (1)
- domain assumption Plant action potentials obey dynamics sufficiently similar to neuronal action potentials that the Hodgkin-Huxley equations remain applicable after modest parameter adjustment.
invented entities (1)
-
POCE and NETO
no independent evidence
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
voltage-dependent rate constants are simplified into constant parameters α and β, thereby reducing the computational load
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Electrical signals as mechanism of photosynthesis regulation in plants,
V . Sukhov, “Electrical signals as mechanism of photosynthesis regulation in plants,”Photosynthesis Research, vol. 130, no. 1-3, pp. 373–387, 2016
work page 2016
-
[2]
A mathematical model of action potential in cells of vascular plants,
V . Sukhov and V . V odeneev, “A mathematical model of action potential in cells of vascular plants,”Journal of Membrane Biology, vol. 232, no. 1-3, p. 59, 2009
work page 2009
-
[3]
Electrical signaling of plants under abiotic stres- sors,
M. Mudrilovet al., “Electrical signaling of plants under abiotic stres- sors,”International Journal of Molecular Sciences, vol. 22, no. 19, p. 10715, 2021
work page 2021
-
[4]
Communication and information theory of single action potential signals in plants,
H. Awan, R. S. Adve, N. Wallbridge, C. Plummer, and A. W. Eckford, “Communication and information theory of single action potential signals in plants,”IEEE Transactions on Nanobioscience, vol. 18, no. 1, pp. 61–73, 2019b
-
[5]
Simulation of action potential propagation in plants,
V . Sukhov, V . Nerush, L. Orlova, and V . V odeneev, “Simulation of action potential propagation in plants,”Journal of Theoretical Biology, vol. 291, pp. 47–55, 2011
work page 2011
-
[6]
The potential of plant action potentials,
J. Lee and P. Calvo, “The potential of plant action potentials,”Synthese, vol. 202, no. 6, 2023
work page 2023
-
[7]
Long-distance plant signaling pathways in response to multiple stressors: the gap in knowledge,
A. E. Huber and T. L. Bauerle, “Long-distance plant signaling pathways in response to multiple stressors: the gap in knowledge,”Journal of Experimental Botany, vol. 67, no. 7, pp. 2063–2079, 2016
work page 2063
-
[8]
Detecting severe plant water stress with machine learning in iot-enabled chamber,
I. Bekkari, S. Lombardo, A. Coviello, and M. Magarini, “Detecting severe plant water stress with machine learning in iot-enabled chamber,” in2024 7th International Balkan Conference on Communications and Networking (BalkanCom). IEEE, 2024
work page 2024
-
[9]
Modelling the role of inter-cellular communication in modulating photosynthesis in plants,
H. Awan, R. S. Adve, N. Wallbridge, C. Plummer, and A. W. Eckford, “Modelling the role of inter-cellular communication in modulating photosynthesis in plants,”IEEE Transactions on Molecular, Biological and Multi-Scale Communications, vol. 1, 2021
work page 2021
- [10]
-
[11]
C. Miaoet al., “Effects of light intensity on growth and quality of lettuce and spinach cultivars in a plant factory,”Plants, vol. 12, no. 18, p. 3337, 2023
work page 2023
-
[12]
S. Proietti, S. Moscatello, F. Riccio, P. Downey, and A. Battistelli, “Con- tinuous lighting promotes plant growth, light conversion efficiency, and nutritional quality of eruca vesicaria (l.) cav. in controlled environment with minor effects due to light quality,”Frontiers in Plant Science, vol. 12, 2021
work page 2021
-
[13]
Light- and dark- induced action potentials in physcomitrella patens,
M. Koselski, K. Trebacz, H. Dziubinska, and E. Krol, “Light- and dark- induced action potentials in physcomitrella patens,”Plant Signaling & Behavior, vol. 3, no. 1, pp. 13–18, 2008
work page 2008
-
[14]
K. Trebacz, W. Simonis, and G. Schonknecht, “Effects of anion channel inhibitors on light-induced potential changes in the liverwort cono- cephalum conicum,”Plant and Cell Physiology, vol. 38, no. 5, pp. 550– 557, 1997
work page 1997
-
[15]
Shade-induced action potentials in helianthus annuus l. originate pri- marily from the epicotyl,
R. Stahlberg, N. R. Stephens, R. E. Cleland, and E. Van V olkenburgh, “Shade-induced action potentials in helianthus annuus l. originate pri- marily from the epicotyl,”Plant Signaling & Behavior, vol. 1, no. 1, pp. 15–22, 2006
work page 2006
-
[16]
J. Yaoet al., “A time-varying parameter estimation method for physi- ological models based on physical information neural networks,”Com- puter Modeling in Engineering & Sciences, vol. 137, no. 3, pp. 2243– 2265, 2023
work page 2023
-
[17]
Light-triggered action potentials in plants,
K. Trebaczet al., “Light-triggered action potentials in plants,”Acta Societatis Botanicorum Poloniae, vol. 58, no. 1, pp. 141–156, 2014
work page 2014
-
[18]
Electrophysiological assessment of plant status outside a faraday cage,
D. Tranet al., “Electrophysiological assessment of plant status outside a faraday cage,”Scientific Reports, vol. 9, no. 1, 2019
work page 2019
-
[19]
J. T. M. Elzenga, H. B. A. Prins, and E. Van V olkenburgh, “Light- induced membrane potential changes of epidermal and mesophyll cells in growing leaves of pisum sativum,”Planta, vol. 197, no. 1, pp. 127– 134, 1995
work page 1995
-
[20]
C. Plieth, B. Sattelmacher, and U. Hansen, “Light-induced cytosolic calcium transients in green plant cells. ll. the effect on a k+ channel as studied by a kinetic analysis in chara corallina,”Planta, vol. 207, no. 1, pp. 52–59, 1998
work page 1998
-
[21]
Model neurons: From hodgkin-huxley to hopfield,
L. F. Abbott and T. B. Kepler, “Model neurons: From hodgkin-huxley to hopfield,” inStatistical Mechanics of Neural Networks. Springer Berlin Heidelberg, s.d., pp. 5–18
-
[22]
A. de Angeli, O. Moran, S. Wege, S. Filleur, G. Ephritikhineet al., “Atp binding to the c terminus of the arabidopsis thaliana nitrate/proton antiporter, atclca, regulates nitrate transport into plant vacuoles,”Journal of Biological Chemistry, vol. 284, no. 39, pp. 26 526–26 532, 2009
work page 2009
-
[23]
Substrate regulation of single potassium and chloride ion channels in arabidopsis plasma membrane,
R. R. Lew, “Substrate regulation of single potassium and chloride ion channels in arabidopsis plasma membrane,”Plant Physiology, vol. 95, no. 2, pp. 642–647, 1991
work page 1991
-
[24]
B. Van Duijn, “Hodgkin-huxley analysis of whole-cell outward rectify- ing k+-currents in protoplasts from tobacco cell suspension cultures,” The Journal of Membrane Biology, vol. 132, no. 1, pp. 77–85, 1993
work page 1993
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.