High-fidelity and Network-based Spatio-temporal Mathematical Models of Alzheimer's Disease Progression and their Validation Against PET-SUVR Imaging Data
Pith reviewed 2026-05-10 03:36 UTC · model grok-4.3
The pith
Three-dimensional models of amyloid-beta and tau spread match PET data more accurately than connectome network models in Alzheimer's disease.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
A high-fidelity biophysical model defined on three-dimensional patient-specific brain geometries reconstructed from MRI provides a more accurate and biologically consistent description of amyloid-beta and tau protein dynamics than a reduced network-based formulation on the brain connectome. Both models are discretized numerically, subjected to sensitivity analysis, and validated against PET-SUVR clinical data using specific tracers for each protein. The three-dimensional approach yields superior results in matching imaging observations despite greater computational demands.
What carries the argument
The high-fidelity three-dimensional patient-specific geometry model for spatio-temporal protein transport and accumulation, compared against the reduced connectome network model.
If this is right
- The 3D model better captures individual variations in disease progression from imaging data.
- Sensitivity analysis reveals which parameters most affect protein concentration patterns.
- The network model offers a faster alternative but may not reliably predict all observed patterns.
- Validation against two different PET tracers confirms the models' applicability to both amyloid-beta and tau proteins.
- Numerical discretizations enable practical simulation of the dynamics on the chosen geometries or networks.
Where Pith is reading between the lines
- If the 3D model proves consistently superior, it could guide development of personalized simulations for predicting disease course in individual patients.
- The framework might extend to modeling other neurodegenerative conditions involving protein misfolding, such as Parkinson's disease.
- Further testing on larger datasets could identify when the cheaper network model is sufficient for clinical research.
- Connecting these models to genetic or lifestyle factors could reveal new ways to intervene in protein accumulation.
Load-bearing premise
The biophysical parameters and brain connectome structure chosen for the models accurately capture the real mechanisms of protein transport and accumulation in the brain.
What would settle it
New PET-SUVR imaging data from a different cohort of Alzheimer's patients where the three-dimensional model's predictions deviate significantly from observed protein distributions while the network model's do not.
Figures












read the original abstract
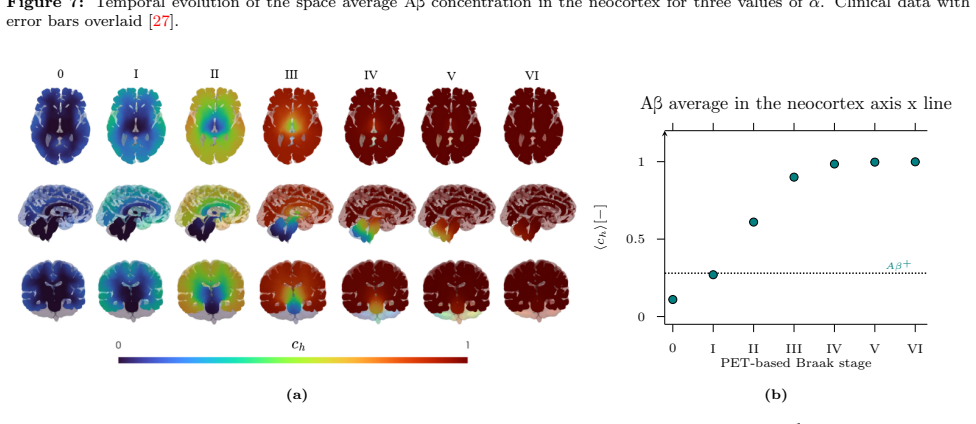
Alzheimer's disease is the most common neurodegenerative disorder. Its pathological development is connected with the misfolding and accumulation of two toxic proteins: amyloid-beta and tau proteins. Mathematical models provide a valuable quantitative tool for monitoring disease progression. In this work, we proposed and compare a novel framework where the spatio-temporal dynamics of amyloid-beta and tau proteins is modeled based on employing either three-dimensional patient-specific geometries or through reduced network-based models defined on the brain connectome. More specifically, a high-fidelity biophysical model is proposed on three-dimensional brain geometries reconstructed from magnetic resonance imaging, whereas a network-based reduced formulation is defined on the brain connectome. For both approaches, a suitable numerical discretisation is proposed. A sensitivity analysis is presented to quantify the influence of model parameters on protein concentration patterns as well as compare the quality of the predictions. For both approaches, the results are validated against PET-SUVR clinical data using 18FAZD4694 for amyloid-beta and 18FMK6240 for tau protein. The results indicate that the three-dimensional model provides the most accurate and biologically consistent description of the disease progression, but remains computationally demanding. On the other hand, the reduced graph-based model is cheaper, but it is not always able to achieve reliable results.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript proposes and compares a high-fidelity three-dimensional biophysical model defined on patient-specific brain geometries reconstructed from MRI with a reduced network-based model on the brain connectome for the spatio-temporal dynamics of amyloid-beta and tau proteins. Both formulations include numerical discretizations, a sensitivity analysis on parameters, and validation of predicted concentration patterns against PET-SUVR data from the 18FAZD4694 tracer for amyloid-beta and the 18FMK6240 tracer for tau. The central conclusion is that the 3D model yields the most accurate and biologically consistent description of disease progression, while the network model is computationally cheaper but less reliably accurate.
Significance. If the superiority of the 3D model is shown to arise from correct representation of protein transport physics rather than fitting flexibility, the work would be significant for computational neuroscience by quantifying the accuracy-efficiency trade-off in Alzheimer's modeling and providing a validated framework for patient-specific simulations. The sensitivity analysis and direct use of two independent PET tracers constitute clear strengths that support reproducibility and parameter influence assessment.
major comments (1)
- [Validation section] Validation section: The claim that the three-dimensional model provides the 'most accurate and biologically consistent description' rests on comparison to the same PET-SUVR datasets used for both models. The manuscript does not demonstrate that the diffusion coefficients, reaction rates, or clearance terms are taken from independent in-vitro or animal studies rather than optimized against these imaging observations; without such external anchoring, the reported advantage of the 3D geometry could arise from its higher spatial degrees of freedom instead of mechanistic fidelity.
minor comments (2)
- [Abstract] The abstract states that 'only two biophysical parameters are used' but does not specify which parameters these are or how their values were selected; this detail should be stated explicitly in the methods to allow readers to assess the fitting procedure.
- Figure captions and the sensitivity analysis presentation would benefit from explicit reporting of error bars or confidence intervals on the PET-SUVR comparison metrics to strengthen the quantitative claims of accuracy.
Simulated Author's Rebuttal
We thank the referee for the detailed and constructive review of our manuscript. The major comment raises an important point about parameter sourcing and the interpretation of model accuracy. We address this directly below and describe the revisions we will make to strengthen the validation section.
read point-by-point responses
-
Referee: [Validation section] Validation section: The claim that the three-dimensional model provides the 'most accurate and biologically consistent description' rests on comparison to the same PET-SUVR datasets used for both models. The manuscript does not demonstrate that the diffusion coefficients, reaction rates, or clearance terms are taken from independent in-vitro or animal studies rather than optimized against these imaging observations; without such external anchoring, the reported advantage of the 3D geometry could arise from its higher spatial degrees of freedom instead of mechanistic fidelity.
Authors: We agree that the manuscript would benefit from greater transparency on this issue. The diffusion, reaction, and clearance parameters were selected from values reported in the prior literature (primarily in-vitro and rodent studies), with limited patient-specific calibration performed only within the ranges supported by those studies. The sensitivity analysis already shows that the 3D model retains its advantage across wide parameter ranges, which argues against the superiority being due solely to extra degrees of freedom. In the revised manuscript we will add an explicit subsection (and accompanying table) that lists every parameter, its literature source, the range explored, and the calibration procedure used for the PET cohorts. We will also qualify the claim of 'most accurate and biologically consistent' to reflect the remaining uncertainty in parameter provenance and note that the network model serves as a lower-dimensional control for overfitting. revision: yes
Circularity Check
No circularity: models and validation rest on external PET-SUVR data and independent biophysical assumptions
full rationale
The paper defines two classes of spatio-temporal models (3D patient-specific PDEs from MRI geometries and reduced network models on the connectome), proposes numerical discretizations, conducts parameter sensitivity analysis, and validates both against independent clinical PET-SUVR datasets (18FAZD4694 for amyloid-beta, 18FMK6240 for tau). No derivation step equates a claimed prediction or first-principles result to its own fitted inputs by construction. The central claim that the 3D model is more accurate rests on quantitative comparison to external imaging observations rather than on self-definition, self-citation chains, or renaming of known patterns. The two biophysical parameters are treated as inputs whose influence is quantified, not as quantities whose values are redefined as outputs.
Axiom & Free-Parameter Ledger
free parameters (2)
- diffusion coefficients for amyloid and tau
- reaction rates and clearance terms
axioms (2)
- domain assumption Protein transport follows reaction-diffusion equations on the given geometry or graph.
- domain assumption The brain connectome accurately represents spatial connectivity for protein propagation.
Reference graph
Works this paper leans on
-
[1]
Neurodegenerative diseases: Expanding the prion concept
L. C. Walker and M. Jucker, “Neurodegenerative diseases: Expanding the prion concept”, Annu. Rev. Neurosci., vol. 38, no. 1, pp. 87–103, 2015
work page 2015
-
[2]
Self-propagation of pathogenic protein aggregates in neurodegenerative diseases
M. Jucker and L. C. Walker, “Self-propagation of pathogenic protein aggregates in neurodegenerative diseases”, Nature, vol. 501, no. 7465, pp. 45–51, 2013
work page 2013
-
[3]
P. Scheltens, B. De Strooper, M. Kivipelto, H. Holstege, G. Ch´ etelat, C. Teunissen, J. Cummings, and W. van der Flier, “Alzheimer’s disease”, The Lancet, vol. 397, pp. 1577–1590, 2021
work page 2021
-
[4]
Amyloid beta: structure, biology and structure-based therapeutic development
G.-F. Chen, T.-H. Xu, Y. Yan, Y. Zhou, Y. Jiang, K. Melcher, and H. E. Xu, “Amyloid beta: structure, biology and structure-based therapeutic development”, Acta Pharmacol. Sin., vol. 38, no. 9, pp. 1205–1235, 2017
work page 2017
-
[5]
M. Jouanne, S. Rault, and A. Voisin-Chiret, “Tau protein aggregation in Alzheimer’s disease: An attractive target for the development of novel therapeutic agents”, Eur. J. Med. Chem., vol. 139, pp. 153–167, 2017
work page 2017
-
[6]
Tau, tangles, and Alzheimer’s disease
L. I. Binder, A. L. Guillozet-Bongaarts, F. Garcia-Sierra, and R. W. Berry, “Tau, tangles, and Alzheimer’s disease”, Biochim. Biophys. Acta, vol. 1739, no. 2–3, pp. 216–223, 2005
work page 2005
-
[7]
Hallmarks of neurodegenerative diseases
D. M. Wilson, M. R. Cookson, L. Van Den Bosch, H. Zetterberg, D. M. Holtzman, and I. Dewachter, “Hallmarks of neurodegenerative diseases”, Cell, vol. 186, no. 4, pp. 693–714, 2023
work page 2023
-
[8]
B. Dubois, C. A. F. von Arnim, N. Burnie, S. Bozeat, and J. Cummings, “Biomarkers in Alzheimer’s disease: Role in early and differential diagnosis and recognition of atypical variants”, Alzheimer’s Res. Ther., vol. 15, p. 175, 2023
work page 2023
-
[9]
Insights into the use of biomarkers in clinical trials in Alzheimer’s disease
T. A. Pascoal, C. S. Aguzzoli, F. Z. Lussier, L. Crivelli, C. K. Suemoto, J. Fortea, P. Rosa-Neto, E. R. Zimmer, P. C. L. Ferreira, and B. Bellavera, “Insights into the use of biomarkers in clinical trials in Alzheimer’s disease”, eBioMedicine, vol. 108, p. 105322, 2024
work page 2024
-
[10]
Core candidate neurochemical and imaging biomarkers of Alzheimer’s disease
H. Hampel, K. B¨ urger, S. J. Teipel, A. L. W. Bokde, H. Zetterberg, and K. Blennow, “Core candidate neurochemical and imaging biomarkers of Alzheimer’s disease”, Alzheimer’s & Dementia, vol. 4, no. 1, pp. 38–48, 2008
work page 2008
-
[11]
PET neuroimaging of Alzheimer’s disease: Radio- tracers and their utility in clinical research
W. Bao, F. Xie, C. Zuo, Y. Guan, and Y. H. Huang, “PET neuroimaging of Alzheimer’s disease: Radio- tracers and their utility in clinical research”, Front. Aging Neurosci., vol. 13, p. 624330, 2021
work page 2021
-
[12]
The spread of prion diseases in the brain: Models of reaction and transport on networks
F. Matth¨ aus, “The spread of prion diseases in the brain: Models of reaction and transport on networks”, J. Biol. Syst., vol. 17, no. 4, pp. 623–641, 2009
work page 2009
-
[13]
The wave of advance of advantageous genes
R. Fisher, “The wave of advance of advantageous genes”, Annals of Eugenics, vol. 7, no. 4, pp. 353–369, 1937
work page 1937
-
[14]
A. N. Kolmogorov, I. G. Petrovskii, and N. S. Piskunov, “Etude de la diffusion avec croissance de la quantit´ e de mati` ere et son application ` a un probl` eme biologique”,Moscow University Mathematics Bulletin, vol. 1, pp. 1–25, 1937. 18
work page 1937
-
[15]
Prion-like spreading of Alzheimer’s disease within the brain’s connectome
S. Fornari, A. Sch¨ afer, M. Jucker, A. Goriely, and E. Kuhl, “Prion-like spreading of Alzheimer’s disease within the brain’s connectome”, J. R. Soc. Interface, vol. 16, no. 159, p. 20190356, 2019
work page 2019
-
[16]
J. Weickenmeier, M. Jucker, A. Goriely, and E. Kuhl, “A physics-based model explains the prion–like fea- tures of neurodegeneration in Alzheimer’s disease, Parkinson’s disease, and amyotrophic lateral sclerosis”, J. Mech. Phys. Solids, vol. 124, pp. 264–281, 2019
work page 2019
-
[17]
The interplay of biochemical and biomechanical degeneration in Alzheimer’s disease
A. Sch¨ afer, J. Weickenmeier, and E. Kuhl, “The interplay of biochemical and biomechanical degeneration in Alzheimer’s disease”, Comput. Methods Appl. Mech. Engrg., vol. 352, pp. 369–388, 2019
work page 2019
-
[18]
M. Corti, F. Bonizzoni, L. Dede’, A. M. Quarteroni, and P. F. Antonietti, “Discontinuous Galerkin methods for Fisher-Kolmogorov equation with application toα-synuclein spreading in Parkinson’s disease”, Comput. Methods Appl. Mech. Engrg., vol. 417, p. 116450, 2023
work page 2023
-
[19]
M. Corti, “Exploring tau protein and amyloid-beta propagation: A sensitivity analysis of mathematical models based on biological data”, Brain Multiphysics, vol. 7, p. 100098, 2023
work page 2023
-
[20]
M. Corti, F. Bonizzoni, and P. F. Antonietti, “Structure preserving polytopal discontinuous Galerkin methods for the numerical modeling of neurodegenerative diseases”, J. Sci. Comput., vol. 100, p. 39, 2024
work page 2024
-
[21]
P. F. Antonietti, M. Corti, S. G´ omez, and I. Perugia, “A structure-preserving ldg discretization of the fisher–kolmogorov equation for modeling neurodegenerative diseases”, Math. Comput. Simul., vol. 241, pp. 351–366, 2026
work page 2026
-
[22]
A structure-preserving discontinuous galerkin scheme for the fisher–kpp equation
F. Bonizzoni, M. Braukhoff, A. J¨ angel, and I. Perugia, “A structure-preserving discontinuous galerkin scheme for the fisher–kpp equation”, Numer. Math., vol. 146, pp. 119–157, 2020
work page 2020
-
[23]
A network diffusion model of disease progression in dementia
A. Raj, A. Kuceyeski, and M. Weiner, “A network diffusion model of disease progression in dementia”, Neuron, vol. 73, no. 6, pp. 1204–1215, 2012
work page 2012
-
[24]
Y. Iturria-Medina, R. C. Sotero, P. J. Toussaint, A. C. Evans, and Alzheimer’s Disease Neuroimaging Ini- tiative, “Epidemic spreading model to characterize misfolded proteins propagation in aging and associated neurodegenerative disorders”, PLoS Comput. Biol., vol. 10, no. 11, p. e1003956, 2014
work page 2014
-
[25]
Bayesian physics-based modeling of tau propagation in Alzheimer’s disease
A. Sch¨ afer, M. Peirlinck, K. Linka, E. Kuhl, and Alzheimer’s Disease Neuroimaging Initiative, “Bayesian physics-based modeling of tau propagation in Alzheimer’s disease”, Front. Physiol., vol. 12, p. 1081, 2021
work page 2021
-
[26]
M. Corti, F. Bonizzoni, P. F. Antonietti, and A. M. Quarteroni, “Uncertainty quantification for Fisher– Kolmogorov equation on graphs with application to patient-specific Alzheimer’s disease”, ESAIM: Math. Model. Numer. Anal., vol. 58, no. 1, pp. 147–176, 2024
work page 2024
-
[27]
Biomarker modeling of Alzheimer’s disease using PET-based Braak staging
J. Therriault, T. A. Pascoal, F. Z. Lussier, C. Tissot, M. Chamoun, G. Bezgin, S. Servaes, A. L. Benedet, N. J. Ashton, T. K. Karikari, J. Lantero-Rodriguez, P. Kunach, Y.-T. Wang, J. Fernandez-Arias, G. Massar- weh, P. Vitali, J.-P. Soucy, P. Saha-Chaudhuri, K. Blennow, H. Zetterberg, S. Gauthier, and P. Rosa-Neto, “Biomarker modeling of Alzheimer’s dise...
work page 2022
-
[28]
H. Braak, I. Alafuzoff, T. Arzberger, H. Kretzschmar, and K. Del Tredici, “Staging of Alzheimer disease- associated neurofibrillary pathology using paraffin sections and immunocytochemistry”,Acta Neuropathol., vol. 112, no. 4, pp. 389–404, 2006
work page 2006
-
[29]
Unified analysis of discontinuous Galerkin methods for elliptic problems
D. Arnold, F. Brezzi, B. Cockburn, and D. Marini, “Unified analysis of discontinuous Galerkin methods for elliptic problems”, SIAM J. Numer. Anal., vol. 39, no. 5, pp. 1749–1779, 2001-02
work page 2001
-
[30]
Generative models for network neuroscience: Prospects and promise
R. F. Betzel and D. S. Bassett, “Generative models for network neuroscience: Prospects and promise”, J. R. Soc. Interface, vol. 14, no. 136, p. 20170623, 2017
work page 2017
-
[31]
Kernels and regularization on graphs
A. J. Smola and R. Kondor, “Kernels and regularization on graphs”, in Learning Theory and Kernel Machines, vol. 2777 of Lecture Notes in Computer Science, pp. 144–158, Springer, 2003
work page 2003
-
[32]
P. J. LaMontagne, T. L. Benzinger, J. C. Morris, S. Keefe, R. Hornbeck, C. Xiong, E. Grant, J. Hassenstab, K. Moulder, A. G. Vlassenko, M. E. Raichle, C. Cruchaga, and D. Marcus, “OASIS-3: Longitudinal neuroimaging, clinical, and cognitive dataset for normal aging and Alzheimer disease”, 2019, MedRxiv. 19
work page 2019
- [33]
-
[34]
R. S. Desikan, F. S´ egonne, B. Fischl, B. T. Quinn, B. C. Dickerson, D. Blacker, R. L. Buckner, A. M. Dale, R. P. Maguire, B. T. Hyman, M. S. Albert, and R. J. Killiany, “An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest”, NeuroImage, vol. 31, no. 3, pp. 968–980, 2006
work page 2006
- [35]
-
[36]
DSI Studio: an integrated tractography platform and fiber data hub for accelerating brain research
F.-C. Yeh, “DSI Studio: an integrated tractography platform and fiber data hub for accelerating brain research”, Nat. Methods, vol. 22, pp. 1617–1619, 2025
work page 2025
-
[37]
The Human Brainnetome Atlas: A new brain atlas based on connectional architecture
L. Fan, H. Li, J. Zhuo, Y. Zhang, J. Wang, L. Chen, Z. Yang, C. Chu, S. Xie, A. R. Laird, P. T. Fox, S. B. Eickhoff, C. Yu, and T. Jiang, “The Human Brainnetome Atlas: A new brain atlas based on connectional architecture”, Cereb. Cortex, vol. 26, no. 8, pp. 3508–3526, 2016
work page 2016
-
[38]
Network diffusion modeling explains longitudinal tau PET data
A. Sch¨ afer, E. C. Mormino, E. Kuhl, and Alzheimer’s Disease Neuroimaging Initiative, “Network diffusion modeling explains longitudinal tau PET data”, Front. Neurosci., vol. 14, p. 566876, 2020
work page 2020
-
[39]
A. Whittington, D. J. Sharp, R. N. Gunn, and Alzheimer’s Disease Neuroimaging Initiative, “Spatiotem- poral distribution ofβ-amyloid in Alzheimer disease is the result of heterogeneous regional carrying capac- ities”, J. Nuc. Med., vol. 59, no. 5, pp. 822–827, 2018
work page 2018
-
[40]
Determining amyloid-βpositivity using [ 18F]AZD4694 PET imaging
J. Therriault, T. A. Pascoal, A. L. Benedet, M. Savard, F. Z. Lussier, C. Tissot, M. Chamoun, S. A. Mathotaarachchi, E. Thomas, S. Gauthier, P. Rosa-Neto, and Alzheimer’s Disease Neuroimaging Initiative, “Determining amyloid-βpositivity using [ 18F]AZD4694 PET imaging”, J. Nucl. Med., vol. 62, no. 2, pp. 247–252, 2021
work page 2021
-
[41]
Longitudinal 18F-MK-6240 tau tangles accumulation follows Braak stages
T. A. Pascoal, A. L. Benedet, D. L. Tudorascu, J. Therriault, S. Mathotaarachchi, M. Savard, F. Z. Lussier, C. Tissot, M. Chamoun, M. S. Kang, J. Stevenson, G. Massarweh, M.-C. Guiot, J.-P. Soucy, S. Gauthier, and P. Rosa-Neto, “Longitudinal 18F-MK-6240 tau tangles accumulation follows Braak stages”, Brain, vol. 144, pp. 3517–3528, 2021
work page 2021
-
[42]
Kinetics of tau aggregation reveals patient–specific tau characteristics among Alzheimer’s cases
T. V. Kamath, N. Klickstein, C. Commins, A. R. Fernandes, D. H. Oakley, M. P. Frosch, B. T. Hyman, and S. Dujardin, “Kinetics of tau aggregation reveals patient–specific tau characteristics among Alzheimer’s cases”, Brain Commun., vol. 3, no. 2, p. fcab096, 2021
work page 2021
-
[43]
Personalised regional modelling predicts tau progression in the human brain
P. Chaggar, J. W. Vogel, A. Pichet Binette, T. B. Thompson, O. Strandberg, N. Mattsson-Carlgren, L. Karlsson, E. Stomrud, S. Jbabdi, S. Magon, G. Klein, O. Hansson, A. Goriely, and Alzheimer’s Disease Neuroimaging Initiative, “Personalised regional modelling predicts tau progression in the human brain”, PLoS Biol., vol. 23, no. 7, p. e3003241, 2025
work page 2025
-
[44]
N. Krishnadas, V. Dor´ e, J. S. Robertson, L. Ward, C. Fowler, C. L. Masters, P. Bourgeat, J. Fripp, V. L. Villemagne, and C. C. Rowe, “Rates of regional tau accumulation in ageing and across the Alzheimer’s disease continuum: An AIBL 18F-MK6240 PET study”, eBioMedicine, vol. 88, p. 104450, 2023
work page 2023
-
[45]
Comparing brain networks of different size and connectivity density using graph theory
B. C. M. van Wijk, C. J. Stam, and A. Daffertshofer, “Comparing brain networks of different size and connectivity density using graph theory”, PLoS ONE, vol. 5, no. 10, 2010
work page 2010
-
[46]
F. Xu, S. Garai, D. Duong-Tran, A. J. Saykin, Y. Zhao, L. Shen, and Alzheimer’s Disease Neuroimaging Initiative, “Consistency of graph theoretical measurements of Alzheimer’s disease fiber density connectomes across multiple parcellation scales”, in Proceedings of the IEEE International Conference on Bioinformatics and Biomedicine (BIBM), pp. 1323–1328, I...
work page 2023
-
[47]
Discontinuous Galerkin approximations of the heterodimer model for protein-protein interaction
P. F. Antonietti, F. Bonizzoni, M. Corti, and A. Dall’Olio, “Discontinuous Galerkin approximations of the heterodimer model for protein-protein interaction”, Comput. Methods Appl. Mech. Engrg., vol. 431, p. 117282, 2024
work page 2024
-
[48]
W. van Oostveen and E. de Lange, “Imaging techniques in Alzheimer’s disease: A review of applications in early diagnosis and longitudinal monitoring”, Int. J. Mol. Sci., vol. 22, no. 4, p. 2110, 2021. 20
work page 2021
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.