Thermodynamic Parametrisation of the Vertebrate Lifetime Cycle Invariant: Biological Proper Time, Allometric Mass-Cancellation, and Clade-Specific Predictions
Pith reviewed 2026-05-07 17:50 UTC · model grok-4.3
The pith
The number of heartbeats in a vertebrate lifetime stays fixed because lifetime entropy production is constant and mass terms cancel under scaling laws.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
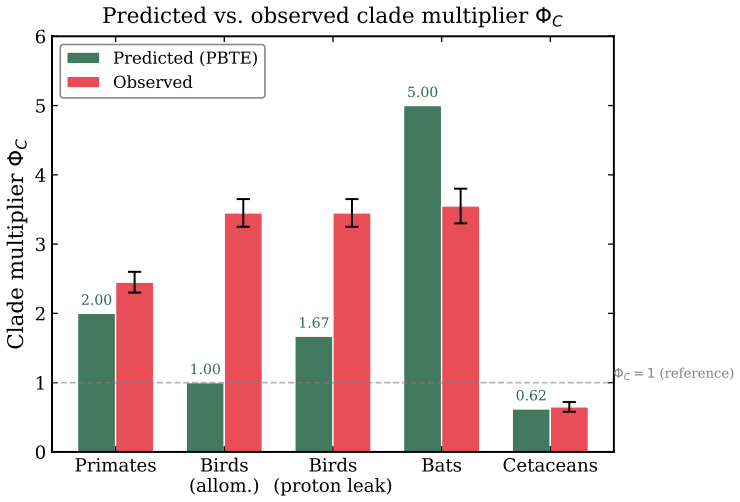
Integration over the lifespan yields lifetime entropy production equal to a constant times the number of cardiac cycles, so the observed constancy of this number corresponds to an approximately constant lifetime entropy budget. Algebraic cancellation under Kleiber and Calder scaling laws removes mass dependence entirely and reproduces the numerical value without free parameters. A multiplicative correction constructed from activity allocation, body temperature, mitochondrial efficiency and extrinsic hazard then predicts long-lived clades as regimes of reduced effective entropy production per cycle.
What carries the argument
The constitutive closure that sets entropy production rate proportional to the intrinsic physiological frequency, integrated over lifespan to give total entropy as constant times total heartbeats.
Where Pith is reading between the lines
- The same scaling cancellation might produce invariants for other repeating physiological processes such as total breaths or total muscle contractions if analogous rate laws hold.
- Interventions that change mitochondrial efficiency or body temperature should shift the effective entropy cost per cycle and thereby alter predicted lifespan in a quantifiable way.
- The framework offers a thermodynamic route to connect lifetime entropy accumulation with cellular damage models of aging without invoking separate mechanisms for each.
Load-bearing premise
Entropy production rate is directly proportional to heart rate as the governing physiological frequency.
What would settle it
Direct calorimetric or biochemical measurements of cumulative entropy production across the full lifespan of vertebrate species spanning a wide mass range that show total entropy varying systematically with body mass instead of remaining constant.
Figures







read the original abstract
Warm-blooded vertebrates accumulate approximately $\Nstar \approx 10^9$ cardiac cycles over a natural lifetime, a striking empirical regularity first quantified by Lindstedt and Calder yet lacking a physical interpretation. We propose that this invariance is consistent with a conserved thermodynamic budget, formulated here as the Principle of Biological Time Equivalence (PBTE). The framework rests on a constitutive closure $\dot{\Sigma} = \sigma_0 f$, which links the entropy production rate to the intrinsic physiological frequency; integration over the lifespan yields $\Sigma_{\mathrm{life}} = \sigma_0 \Nstar$, so that the observed constancy of $\Nstar$ corresponds to an approximately constant lifetime entropy budget. Algebraic exponent cancellation under Kleiber and Calder scaling laws, $\sigstar \propto M^{3/4+1/4-1}=M^0$, is consistent with mass-independence and reproduces the numerical value $N_0 \approx 1.52\times10^9$ without free parameters. The framework offers a thermodynamically consistent account of two outstanding problems: the origin of the numerical value of $\Nstar$ and the systematic deviations observed across clades. A multiplicative correction factor $\Phi_C$, constructed from physiological determinants -- activity allocation, body temperature, mitochondrial efficiency, and extrinsic hazard -- predicts long-lived clades as regimes of reduced effective entropy production per cardiac cycle.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript proposes that the empirical invariance N* ≈ 10^9 cardiac cycles over vertebrate lifetimes reflects a conserved lifetime entropy budget Σ_life = σ0 N*, formalized via the Principle of Biological Time Equivalence (PBTE) and the constitutive closure Σ̇ = σ0 f. Integration of this relation, combined with Kleiber (BMR ∝ M^{3/4}) and Calder (f ∝ M^{-1/4}) scaling, yields mass-independent σ* through algebraic cancellation (σ* ∝ M^{3/4 + 1/4 - 1} = M^0) and reproduces the numerical value N0 ≈ 1.52 × 10^9 without free parameters. A multiplicative correction Φ_C incorporating activity, temperature, mitochondrial efficiency, and hazard is introduced to account for clade-specific deviations.
Significance. If the central relation holds, the work supplies a thermodynamic interpretation for a well-known biological scaling regularity and generates falsifiable clade predictions. The algebraic cancellation itself is a clean consistency check on existing allometric laws, but the absolute scale and the entropy-budget interpretation rest entirely on the posited closure.
major comments (2)
- [Abstract / §2] Abstract and §2 (constitutive closure): The relation Σ̇ = σ0 f is introduced as an axiom without derivation from statistical mechanics, measured heat fluxes, or mitochondrial irreversibility. This closure is load-bearing for the claim that constancy of N* implies constancy of Σ_life; absent a first-principles or empirical justification, the thermodynamic budget interpretation remains conjectural rather than derived.
- [Abstract] Abstract (numerical value): The reproduction of N0 ≈ 1.52 × 10^9 is stated to be parameter-free, yet it necessarily incorporates the numerical prefactors already present in the cited Kleiber and Calder relations. The manuscript does not demonstrate that σ0 is independently fixed by thermodynamics; the absolute scale therefore inherits calibration from prior empirical fits.
minor comments (1)
- [Abstract] Notation: The symbol σ* is used both for the mass-specific entropy production rate and for its lifetime-integrated value; a clearer distinction (e.g., σ̇* vs. Σ*) would improve readability.
Simulated Author's Rebuttal
We appreciate the referee's thorough review and valuable feedback on our manuscript. The comments highlight important aspects of the constitutive closure and the parameter-free nature of the numerical prediction. We agree that additional clarification is needed for the former and will revise the text accordingly. For the latter, we provide further explanation while maintaining the core claim. Our point-by-point responses follow.
read point-by-point responses
-
Referee: [Abstract / §2] Abstract and §2 (constitutive closure): The relation Σ̇ = σ0 f is introduced as an axiom without derivation from statistical mechanics, measured heat fluxes, or mitochondrial irreversibility. This closure is load-bearing for the claim that constancy of N* implies constancy of Σ_life; absent a first-principles or empirical justification, the thermodynamic budget interpretation remains conjectural rather than derived.
Authors: We concur that the constitutive relation dot{Σ} = σ0 f is posited as a foundational closure rather than derived from microscopic statistical mechanics or direct measurements of mitochondrial irreversibility. It is motivated by the principle that the dominant source of entropy production in homeothermic vertebrates is tied to the metabolic rate, which in turn scales with the frequency of cardiac cycles as the fundamental physiological timer. Although we do not provide a first-principles derivation, the relation is consistent with empirical allometric observations linking heat production to heart rate. In the revised version, we will expand the discussion in §2 to include references to thermodynamic analyses of biological systems and empirical estimates of entropy production per cycle from literature on metabolic efficiency. We will also explicitly state the phenomenological status of the PBTE to avoid overstating its foundational character. revision: yes
-
Referee: [Abstract] Abstract (numerical value): The reproduction of N0 ≈ 1.52 × 10^9 is stated to be parameter-free, yet it necessarily incorporates the numerical prefactors already present in the cited Kleiber and Calder relations. The manuscript does not demonstrate that σ0 is independently fixed by thermodynamics; the absolute scale therefore inherits calibration from prior empirical fits.
Authors: The referee correctly notes that the specific numerical value arises from combining the prefactors in the Kleiber scaling for basal metabolic rate and the Calder scaling for heart rate, together with the value of σ0. However, the manuscript's claim of being 'without free parameters' refers to the absence of any new adjustable constants introduced to fit the invariance; the mass independence follows directly from the exponent cancellation (3/4 + 1/4 - 1 = 0), and the resulting constant is determined solely by the established allometric coefficients and the universal σ0. We do not claim that σ0 is fixed by pure thermodynamics independent of data; rather, the framework shows that once σ0 is set by the typical lifetime entropy budget, the invariance holds across masses. We will revise the abstract and relevant sections to clarify this distinction and acknowledge the empirical origin of the absolute scale. revision: partial
Circularity Check
Empirical scaling laws and posited constitutive closure re-express known N* constancy as thermodynamic budget
specific steps
-
fitted input called prediction
[Abstract]
"Algebraic exponent cancellation under Kleiber and Calder scaling laws, σ* ∝ M^{3/4+1/4-1}=M^0, is consistent with mass-independence and reproduces the numerical value N0 ≈ 1.52×10^9 without free parameters."
Mass independence is obtained by direct addition of the empirical exponents (3/4 + 1/4 − 1 = 0). The specific numerical value N0 is recovered only after inserting the numerical prefactors and constants already determined by fitting those same scaling laws to biological data; the result is therefore a re-calculation of the input empirical regularity, not a parameter-free thermodynamic derivation.
-
other
[Abstract]
"The framework rests on a constitutive closure Σ̇ = σ0 f, which links the entropy production rate to the intrinsic physiological frequency; integration over the lifespan yields Σ_life = σ0 N*, so that the observed constancy of N* corresponds to an approximately constant lifetime entropy budget."
The equality Σ_life = σ0 N* and the interpretation of constant N* as constant entropy budget are obtained solely by assuming the proportionality Σ̇ = σ0 f. No section derives this closure from measured entropy fluxes or statistical mechanics; the budget constancy is therefore definitionally equivalent to the posited relation rather than an independent consequence.
full rationale
The derivation posits the closure Σ̇ = σ0 f without first-principles justification, integrates to Σ_life = σ0 N*, then invokes Kleiber/Calder scalings to obtain mass cancellation by exponent arithmetic and to recover the numerical N0 value. Because both the absolute scale of N0 and the constancy interpretation depend on prefactors and constants taken from the same empirical scaling relations (plus the un-derived proportionality to heart rate), the central claims reduce to re-labeling of fitted inputs rather than an independent thermodynamic prediction. The framework remains consistent with data but does not derive the invariant from statistical mechanics.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption Kleiber's law for metabolic scaling (metabolic rate ∝ M^{3/4})
- domain assumption Calder scaling laws relating heart rate and lifespan to mass
invented entities (3)
-
Principle of Biological Time Equivalence (PBTE)
no independent evidence
-
Constitutive closure Σ̇ = σ0 f
no independent evidence
-
Multiplicative correction factor Φ_C
no independent evidence
Reference graph
Works this paper leans on
-
[1]
Rubner,Das Problem der Lebensdauer(Oldenbourg, Munich, 1908)
M. Rubner,Das Problem der Lebensdauer(Oldenbourg, Munich, 1908)
work page 1908
-
[2]
S. L. Lindstedt and W. A. Calder, Q. Rev. Biol.56, 1 (1981)
work page 1981
-
[3]
S. D. Livingstone and L. A. Kuehn, Aviat. Space Environ. Med.50, 1037 (1979)
work page 1979
-
[4]
H. J. Levine, J. Am. Coll. Cardiol.30, 1104 (1997). 49
work page 1997
-
[5]
M. A. Taye, J. R. Soc. Interface (submitted 2026). [The empirical lifetime cycle invariant of endothermic vertebrates: a 230-species comparative dataset, statistical validation, and falsifi- ability criteria.] The complete species dataset underlying all statistical results cited from this reference is reproduced in Appendix B of the present paper and can ...
work page 2026
-
[6]
G. B. West, J. H. Brown, and B. J. Enquist, Science276, 122 (1997)
work page 1997
-
[7]
D. S. Glazier, Proc. R. Soc. B289, 20221605 (2022)
work page 2022
-
[8]
Pearl,The Rate of Living(Knopf, New York, 1928)
R. Pearl,The Rate of Living(Knopf, New York, 1928)
work page 1928
-
[9]
J. R. Speakman, J. Exp. Biol.208, 1717 (2005)
work page 2005
-
[10]
Prigogine,Introduction to Thermodynamics of Irreversible Processes, 3rd ed
I. Prigogine,Introduction to Thermodynamics of Irreversible Processes, 3rd ed. (Wiley- Interscience, New York, 1967)
work page 1967
- [11]
-
[12]
Schr¨ odinger,What Is Life?(Cambridge University Press, Cambridge, 1944)
E. Schr¨ odinger,What Is Life?(Cambridge University Press, Cambridge, 1944)
work page 1944
- [13]
-
[14]
W. A. Calder,Size, Function, and Life History(Harvard University Press, Cambridge, MA, 1984)
work page 1984
- [15]
-
[16]
A. J. Hulbertet al., Physiol. Rev.87, 1175 (2007)
work page 2007
-
[17]
R. J. Colmanet al., Nat. Commun.5, 3557 (2014)
work page 2014
-
[18]
Herculano-Houzel, PLOS ONE6, e17514 (2011)
S. Herculano-Houzel, PLOS ONE6, e17514 (2011)
work page 2011
-
[19]
A. K. Yegianet al., Proc. Natl Acad. Sci. USA121, e2313703121 (2024)
work page 2024
- [20]
-
[21]
G. S. Wilkinson and J. M. South, Aging Cell1, 124 (2002)
work page 2002
- [22]
-
[23]
C. E. Ogburnet al., J. Gerontol. A56, B468 (2001)
work page 2001
-
[24]
S. R. Noren and T. M. Williams, J. Exp. Biol.203, 3601 (2000)
work page 2000
- [25]
- [26]
- [27]
-
[28]
Human Ageing Genomic Resources,AnAge build 15(2023),https://genomics.senescence. info/species/. 50
work page 2023
-
[29]
R. Prinzinger, A. Pr¨ asmar, and E. Schleucher, Comp. Biochem. Physiol. A99, 499 (1991)
work page 1991
-
[30]
M. D. Brandet al., Biochem. J.392, 353 (2000)
work page 2000
-
[31]
J. A. Goldbogenet al., Proc. Natl Acad. Sci. USA116, 25329 (2019)
work page 2019
-
[32]
T. M. Williamset al., Nat. Commun.6, 6055 (2015)
work page 2015
- [33]
-
[34]
O. R. P. Bininda-Emondset al., Nature446, 507 (2007)
work page 2007
-
[35]
J. F. Gilloolyet al., Science293, 2248 (2001)
work page 2001
-
[36]
K. E. Joneset al., Ecology90, 2648 (2009)
work page 2009
-
[37]
C. P. Lymanet al.,Hibernation and Torpor in Mammals and Birds(Academic Press, New York, 1982)
work page 1982
-
[38]
Uetzet al.,The Reptile Database(2023),http://www.reptile-database.org/
P. Uetzet al.,The Reptile Database(2023),http://www.reptile-database.org/
work page 2023
-
[39]
K. A. Christian and B. W. Weavers, Copeia1999, 688 (1999)
work page 1999
- [40]
-
[41]
(2026)The Principle of Biological Time Equivalence
Taye, M.A. (2026)The Principle of Biological Time Equivalence. West Los Angeles College [in preparation]
work page 2026
-
[42]
(2026) Neural investment as an entropy-budget strategy.J
Taye, M.A. (2026) Neural investment as an entropy-budget strategy.J. Theor. Biol.[submit- ted]
work page 2026
-
[43]
(2026) Three physiological strategies, one invariant.Proc
Taye, M.A. (2026) Three physiological strategies, one invariant.Proc. R. Soc. B[submitted]
work page 2026
-
[44]
Ruf, T. & Geiser, F. (2015) Daily torpor and hibernation.Biol. Rev.90, 891–926
work page 2015
-
[45]
(1956) On the origin of cancer cells.Science123, 309–314
Warburg, O. (1956) On the origin of cancer cells.Science123, 309–314. Appendix A. Detailed Derivation of the Entropy Cost per Beat and the Cycle-Count Scaling Law This appendix gives a detailed derivation of the entropy-per-beat representation, the lifetime cycle- count relation, and the power-law dependence of lifetime cardiac cycles on the control param...
work page 1956
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.