Homology-based Morphometry of Brain Atrophy: Methods and Applications
Pith reviewed 2026-05-07 17:26 UTC · model grok-4.3
The pith
Persistent homology of T1 MRI yields topological biomarkers that quantify brain atrophy and separate Alzheimer's patients from controls without nonlinear registration.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The central claim is that persistent homology, via slice-wise Euclidean distance transforms on tissue masks and alpha-filtrations on full scans, extracts multiscale topological features that quantify thinning, sulcal widening, and ventricular enlargement; these features distinguish Alzheimer's disease from cognitively normal participants at ROC-AUC 0.895 in cross-sectional analysis and detect greater short-interval structural change in Alzheimer's subjects than in controls in longitudinal data, all without nonlinear registration to a template.
What carries the argument
Two complementary pipelines based on persistent homology: Pipeline 1 applies the Euclidean distance transform to tissue masks in a slice-wise manner to quantify regional thinning; Pipeline 2 uses alpha-filtrations to compute structural similarity between scans and thereby measure disease-related geometric divergence.
If this is right
- Pipeline 1 supports between-subject group comparisons with peak localization to medial temporal regions using only T1 MRI.
- Pipeline 2 supports within-subject longitudinal designs in which follow-up scans remain closest to their own baselines.
- Alzheimer's subjects exhibit measurably greater short-interval topological change than cognitively normal subjects.
- The two pipelines together supply interpretable topological biomarkers usable for both cross-sectional and longitudinal studies without template normalization.
Where Pith is reading between the lines
- The same distance-transform and alpha-filtration approach could be tested on other conditions that produce focal or diffuse tissue loss where template alignment is unreliable.
- Combining the two pipelines in a single analysis workflow might allow simultaneous cross-sectional classification and longitudinal tracking within one study.
- Validation on multi-site data with known variation in acquisition parameters would test whether the features remain stable across different scanners.
Load-bearing premise
The extracted topological features from distance transforms and alpha-filtrations specifically reflect disease-related atrophy rather than scan quality, preprocessing choices, or unrelated geometric variations.
What would settle it
A dataset containing only scanner or preprocessing differences with no induced atrophy, yet producing separation or change scores comparable to those seen between Alzheimer's and control groups, would indicate the features are not specific to atrophy.
Figures
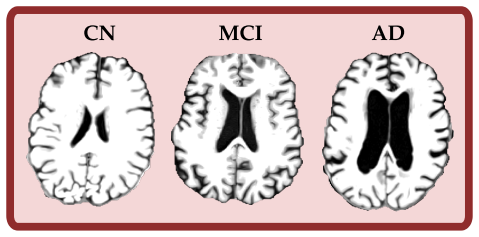








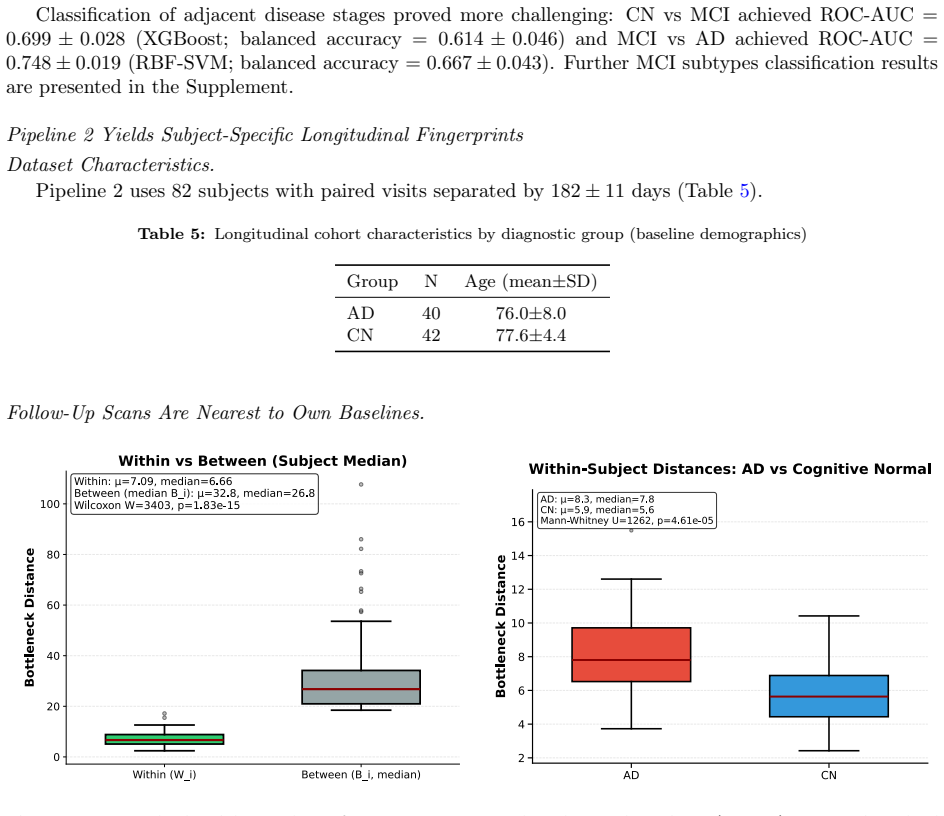







read the original abstract
Understanding the structure of the brain, and how it changes with time and disease, is a core goal of structural neuroimaging. Contemporary approaches to structural brain analysis are dominated by voxel-wise, mass-univariate methods such as voxel-based morphometry (VBM). However, these techniques require images to be normalized to a standard template, which can obscure subject-specific geometric features. Normalization to a common stereotactic space can also be problematic when comparing groups with substantial brain pathology, lesions, or other anatomical abnormalities. Here, we introduce two complementary pipelines based on persistent homology (PH), a tool from topological data analysis, to quantify multiscale geometric features of structural T1-weighted MRI scans. Pipeline 1 quantifies regional thinning by applying the Euclidean distance transform to tissue masks in a slice-wise manner. Pipeline 2 uses \(\alpha\)-filtrations to measure structural similarity between pairs of scans, capturing sulcal widening and ventricular enlargement. Synthetic experiments with controlled induced lesions showed that Pipeline 1 is best suited to between-subject analyses, whereas Pipeline 2 is better suited to within-subject designs. Applied to real-world data from the Alzheimer's Disease Neuroimaging Initiative (ADNI), Pipeline 1 separated Alzheimer's disease (AD) from cognitively normal (CN) participants using single-modality T1-weighted MRI without nonlinear registration (ROC-AUC = 0.895), with peak effects localized to medial temporal regions. Pipeline 2 captured disease-related longitudinal change, with follow-up scans remaining closest to their own baselines and AD subjects showing greater short-interval change than CN subjects. Together, these pipelines provide interpretable topological biomarkers for cross-sectional group comparisons and longitudinal tracking.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper introduces two persistent homology pipelines for structural T1-weighted MRI analysis without nonlinear registration. Pipeline 1 applies slice-wise Euclidean distance transforms to tissue masks to quantify regional thinning, achieving ROC-AUC 0.895 for AD vs CN separation on ADNI data with peak effects in medial temporal regions. Pipeline 2 uses alpha-filtrations for pairwise structural similarity to track longitudinal change, with AD subjects showing greater short-interval differences than CN. Synthetic lesion experiments indicate Pipeline 1 suits between-subject and Pipeline 2 within-subject designs.
Significance. If the performance and localization claims hold after rigorous confound control, the work offers template-free topological biomarkers that preserve subject-specific geometry, addressing limitations of VBM in pathological brains. The dual pipelines for cross-sectional classification and longitudinal tracking, supported by synthetic validation, could provide interpretable alternatives for Alzheimer's morphometry research.
major comments (3)
- [Abstract] Abstract: The reported ROC-AUC = 0.895 for Pipeline 1 is presented without error bars, details on the classifier or feature aggregation, cross-validation protocol, or statistical significance testing; this is load-bearing for the central claim of effective separation and localization to medial temporal regions.
- [Synthetic experiments] Synthetic experiments section: Controlled lesion experiments demonstrate sensitivity to induced changes but do not replicate ADNI confounds (scanner variability, motion, contrast differences); without explicit regression of ICV, age, or scanner effects in the real-data pipeline, the AUC cannot be confidently attributed to disease-specific atrophy rather than global geometry or preprocessing artifacts.
- [Results] Results, localization paragraph: Peak effects localized to medial temporal regions presupposes a regional mapping from slice-wise 2D distance transform features, but the absence of nonlinear registration leaves unclear how spatial correspondence across subjects is achieved without introducing implicit alignment or global shape biases.
minor comments (2)
- [Methods] The alpha-filtration notation should be rendered consistently (e.g., as α-filtrations) in text, equations, and figure captions.
- [Introduction] The introduction would benefit from additional citations to existing persistent homology applications in neuroimaging to better contextualize the novelty of the two pipelines.
Simulated Author's Rebuttal
We thank the referee for the detailed and constructive review. We address each major comment below with clarifications from the manuscript and proposed revisions to improve clarity and rigor.
read point-by-point responses
-
Referee: [Abstract] Abstract: The reported ROC-AUC = 0.895 for Pipeline 1 is presented without error bars, details on the classifier or feature aggregation, cross-validation protocol, or statistical significance testing; this is load-bearing for the central claim of effective separation and localization to medial temporal regions.
Authors: The abstract summarizes the key result while the methods and results sections provide the supporting details: features are aggregated from slice-wise persistent homology barcodes of tissue distance transforms, classified via support vector machine with 10-fold cross-validation, and significance assessed via permutation testing. We agree the abstract would benefit from more context for this central metric. In revision we will add the AUC with bootstrapped 95% CI, a brief note on the classifier and CV scheme, and reference to the full protocol in the main text. revision: yes
-
Referee: [Synthetic experiments] Synthetic experiments section: Controlled lesion experiments demonstrate sensitivity to induced changes but do not replicate ADNI confounds (scanner variability, motion, contrast differences); without explicit regression of ICV, age, or scanner effects in the real-data pipeline, the AUC cannot be confidently attributed to disease-specific atrophy rather than global geometry or preprocessing artifacts.
Authors: The synthetic lesions were designed to isolate topological sensitivity to focal atrophy in a controlled manner, complementing rather than replacing real-data validation. We acknowledge that they do not capture all scanner or motion confounds. In the revised manuscript we will add explicit linear regression of ICV, age, and scanner site as covariates prior to classification, report the adjusted performance metrics, and include a dedicated paragraph discussing residual confounds and their potential impact on interpretation. revision: yes
-
Referee: [Results] Results, localization paragraph: Peak effects localized to medial temporal regions presupposes a regional mapping from slice-wise 2D distance transform features, but the absence of nonlinear registration leaves unclear how spatial correspondence across subjects is achieved without introducing implicit alignment or global shape biases.
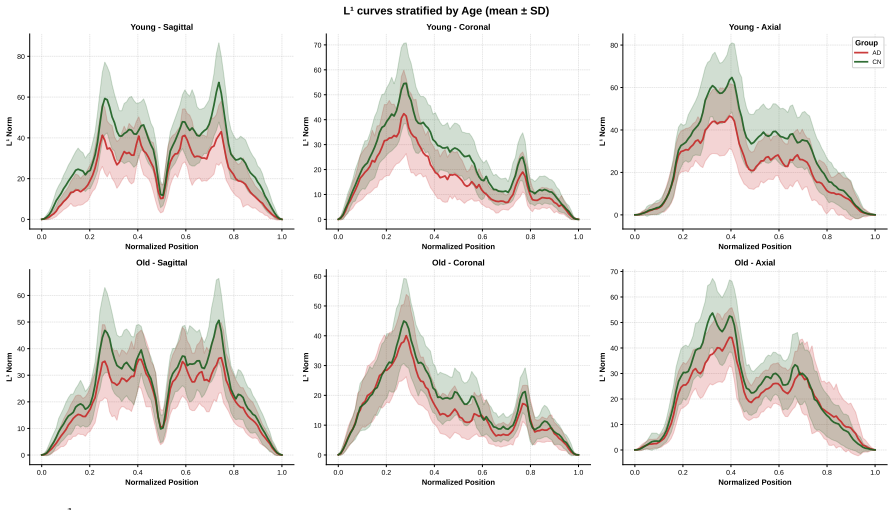
Authors: Regional localization is performed by mapping slices to anatomical territories using subject-native landmarks (AC-PC alignment and standard slice positioning relative to the temporal poles) without nonlinear warping; features from slices corresponding to the medial temporal lobe are then aggregated. This approach deliberately avoids template-induced distortion. We will expand the methods to describe the exact slice-selection criteria and add a supplementary figure showing the regional feature masks overlaid on example native-space images to make the correspondence explicit and address concerns about implicit biases. revision: partial
Circularity Check
No significant circularity; results are empirical applications of topological pipelines
full rationale
The paper presents two pipelines using persistent homology (slice-wise Euclidean distance transforms and alpha-filtrations) applied to T1 MRI tissue masks. The key reported result (ROC-AUC 0.895 separating AD from CN) is an empirical classification outcome on ADNI data, not a mathematical derivation or prediction reduced to fitted parameters. No equations, self-definitional steps, or load-bearing self-citations appear that would make any claim equivalent to its inputs by construction. Synthetic lesion controls and longitudinal tracking are described as validation experiments rather than tautological outputs. The work is therefore self-contained against external data benchmarks.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Persistent homology applied to Euclidean distance transforms and alpha-filtrations of tissue masks captures multiscale geometric features relevant to atrophy.
Reference graph
Works this paper leans on
-
[1]
I. Sebenius, L. Dorfschmidt, J. Seidlitz, A. Alexander-Bloch, S. E. Morgan, E. Bullmore, Structural mri of brain similarity networks, Nature Reviews Neuroscience 26 (1) (2024) 42–59.doi:10.1038/ s41583-024-00882-2. URLhttp://dx.doi.org/10.1038/s41583-024-00882-2
-
[2]
J. Ashburner, K. Friston, Voxel-based morphometry—the methods 11 (6) (2000) 805–821.doi:10. 1006/nimg.2000.0582. URLhttps://www.sciencedirect.com/science/article/pii/S105381190090582X
-
[3]
S. Genon, S. B. Eickhoff, S. Kharabian, Linking interindividual variability in brain structure to be- haviour, Nature Reviews Neuroscience 23 (5) (2022) 307–318.doi:10.1038/s41583-022-00584-7. URLhttp://dx.doi.org/10.1038/s41583-022-00584-7
-
[4]
D. Liloia, D. A. Zamfira, M. Tanaka, J. Manuello, A. Crocetta, R. Keller, M. Cozzolino, S. Duca, F. Cauda, T. Costa, Disentangling the role of gray matter volume and concentration in autism spectrum disorder: A meta-analytic investigation of 25 years of voxel-based morphometry research, Neuroscience and Biobehavioral Reviews 164 (2024) 105791.doi:10.1016/...
-
[5]
L. K. Ferreira, B. S. Diniz, O. V. Forlenza, G. F. Busatto, M. V. Zanetti, Neurostructural predictors of alzheimer’s disease: A meta-analysis of vbm studies, Neurobiology of Aging 32 (10) (2011) 1733–1741. doi:10.1016/j.neurobiolaging.2009.11.008. URLhttp://dx.doi.org/10.1016/j.neurobiolaging.2009.11.008
-
[6]
J. Rowe, L. Hughes, C. Williams-Gray, S. Bishop, S. Fallon, R. Barker, A. Owen, The val158met comt polymorphism’s effect on atrophy in healthy aging and parkinson’s disease, Neurobiology of Aging 31 (6) (2010) 1064–1068.doi:10.1016/j.neurobiolaging.2008.07.009. URLhttp://dx.doi.org/10.1016/j.neurobiolaging.2008.07.009
-
[7]
S. N. Yaakub, R. A. Heckemann, S. S. Keller, C. J. McGinnity, B. Weber, A. Hammers, On brain atlas choice and automatic segmentation methods: a comparison of maper and freesurfer using three atlas databases, Scientific Reports 10 (1) (Feb. 2020).doi:10.1038/s41598-020-57951-6. URLhttp://dx.doi.org/10.1038/s41598-020-57951-6
-
[8]
L. Z. Diaz-de Grenu, J. Acosta-Cabronero, Y. F. V. Chong, J. M. Pereira, S. A. Sajjadi, G. B. Williams, P. J. Nestor, A brief history of voxel-based grey matter analysis in alzheimer’s disease, Journal of Alzheimer’s Disease 38 (3) (2013) 647–659.doi:10.3233/jad-130362. URLhttp://dx.doi.org/10.3233/JAD-130362
-
[9]
X. Zhou, R. Wu, Y. Zeng, Z. Qi, S. Ferraro, L. Xu, X. Zheng, J. Li, M. Fu, S. Yao, K. M. Kendrick, B. Becker, Choice of voxel-based morphometry processing pipeline drives variability in the location of neuroanatomical brain markers, Communications Biology 5 (1) (Sep. 2022).doi: 10.1038/s42003-022-03880-1. URLhttp://dx.doi.org/10.1038/s42003-022-03880-1
-
[10]
N. R. Giuliani, V. D. Calhoun, G. D. Pearlson, A. Francis, R. W. Buchanan, Voxel-based morphom- etry versus region of interest: a comparison of two methods for analyzing gray matter differences in schizophrenia, Schizophrenia Research 74 (2–3) (2005) 135–147.doi:10.1016/j.schres.2004.08.019. URLhttp://dx.doi.org/10.1016/j.schres.2004.08.019
-
[11]
S. Henley, G. Ridgway, R. Scahill, S. Klöppel, S. Tabrizi, N. Fox, J. Kassubek, Pitfalls in the use of voxel-based morphometry as a biomarker: Examples from huntington disease, American Journal of Neuroradiology 31 (4) (2009) 711–719.doi:10.3174/ajnr.a1939. URLhttp://dx.doi.org/10.3174/ajnr.A1939 17
-
[12]
A. Eklund, T. E. Nichols, H. Knutsson, Cluster failure: Why fmri inferences for spatial extent have inflatedfalse-positiverates, ProceedingsoftheNationalAcademyofSciences113(28)(2016)7900–7905. doi:10.1073/pnas.1602413113. URLhttp://dx.doi.org/10.1073/pnas.1602413113
-
[13]
C. Scarpazza, G. Sartori, M. De Simone, A. Mechelli, When the single matters more than the group: Very high false positive rates in single case voxel based morphometry, NeuroImage 70 (2013) 175–188. doi:10.1016/j.neuroimage.2012.12.045. URLhttp://dx.doi.org/10.1016/j.neuroimage.2012.12.045
-
[14]
J. Wen, E. Thibeau-Sutre, M. Diaz-Melo, J. Samper-González, A. Routier, S. Bottani, D. Dormont, S. Durrleman, N. Burgos, O. Colliot, Convolutional neural networks for classification of alzheimer’s disease: Overview and reproducible evaluation, Medical Image Analysis 63 (2020) 101694.doi:10. 1016/j.media.2020.101694. URLhttps://www.sciencedirect.com/scienc...
-
[15]
M. A. Ebrahimighahnavieh, S. Luo, R. Chiong, Deep learning to detect alzheimer’s disease from neu- roimaging: A systematic literature review, Computer Methods and Programs in Biomedicine 187 (2020) 105242.doi:10.1016/j.cmpb.2019.105242. URLhttps://www.sciencedirect.com/science/article/pii/S0169260719309318
-
[16]
Y. Singh, C. Farrelly, Q. Hathaway, et al., Topological data analysis in medical imaging: Current state of the art 14 (2023) 58.doi:10.1186/s13244-023-01413-w. URLhttps://insightsimaging.springeropen.com/articles/10.1186/s13244-023-01413-w
-
[17]
A. Salch, A. Regalski, H. Abdallah, R. Suryadevara, M. J. Catanzaro, V. A. Diwadkar, From math- ematics to medicine: A practical primer on topological data analysis (tda) and the development of related analytic tools for the functional discovery of latent structure in fmri data, PLOS ONE 16 (8) (2021) e0255859.doi:10.1371/journal.pone.0255859. URLhttp://d...
-
[18]
S. G. De Benedictis, G. Gargano, G. Settembre, Enhanced mri brain tumor detection and classification via topological data analysis and low-rank tensor decomposition, Journal of Computational Mathemat- ics and Data Science 13 (2024) 100103.doi:10.1016/j.jcmds.2024.100103. URLhttp://dx.doi.org/10.1016/j.jcmds.2024.100103
-
[19]
A. A. Olave, J. A. Perea, F. Gómez, Revealing brain network dynamics during the emotional state of suspense using tda, Network Neuroscience 9 (4) (2025) 1352–1376.doi:10.1162/netn.a.34. URLhttp://dx.doi.org/10.1162/NETN.a.34
-
[20]
D. Cohen-Steiner, H. Edelsbrunner, J. Harer, Stability of persistence diagrams 37 (1) (2007) 103–120. doi:10.1007/s00454-006-1276-5. URLhttps://link.springer.com/article/10.1007/s00454-006-1276-5
-
[21]
H. Edelsbrunner, J. Harer, Computational Topology: An Introduction, American Mathematical Society, 2010
work page 2010
-
[22]
A. François, R. Tinarrage, Train-free segmentation in mri with cubical persistent homology (2024). arXiv:2401.01160,doi:10.48550/arXiv.2401.01160. URLhttps://arxiv.org/abs/2401.01160
work page internal anchor Pith review doi:10.48550/arxiv.2401.01160 2024
-
[23]
L. Crawford, A. Monod, A. X. Chen, S. Mukherjee, R. Rabadán, Predicting clinical outcomes in glioblastoma: An application of topological and functional data analysis, Journal of the American Statistical Association 115 (531) (2020) 1139–1150.doi:10.1080/01621459.2019.1671198. 18
-
[24]
M. K. Chung, P. Bubenik, P. T. Kim, Persistence Diagrams of Cortical Surface Data, Springer Berlin Heidelberg, 2009, p. 386–397.doi:10.1007/978-3-642-02498-6_32. URLhttp://dx.doi.org/10.1007/978-3-642-02498-6_32
-
[25]
M. K. Chung, J. L. Hanson, J. Ye, R. J. Davidson, S. D. Pollak, Persistent homology in sparse regres- sion and its application to brain morphometry, IEEE Transactions on Medical Imaging 34 (9) (2015) 1928–1939.doi:10.1109/tmi.2015.2416271. URLhttp://dx.doi.org/10.1109/TMI.2015.2416271
-
[26]
M. K. Chung, T. Azizi, J. L. Hanson, A. L. Alexander, S. D. Pollak, R. J. Davidson, Altered topological structure of the brain white matter in maltreated children through topological data analysis, Network Neuroscience 8 (1) (2024) 355–376.doi:10.1162/netn_a_00355. URLhttp://dx.doi.org/10.1162/netn_a_00355
-
[27]
J. S. Phillips, N. Adluru, M. K. Chung, H. Radhakrishnan, C. A. Olm, P. A. Cook, J. C. Gee, K. A. Q. Cousins, S. Arezoumandan, D. A. Wolk, C. T. McMillan, M. Grossman, D. J. Irwin, Greater white matter degeneration and lower structural connectivity in non-amnestic vs. amnestic alzheimer’s disease, Frontiers in Neuroscience 18 (Mar. 2024).doi:10.3389/fnins...
-
[28]
D. Pachauri, C. Hinrichs, M. K. Chung, S. C. Johnson, V. Singh, Topology-based kernels with applica- tion to inference problems in alzheimer’s disease, IEEE Transactions on Medical Imaging 30 (10) (2011) 1760–1770.doi:10.1109/TMI.2011.2147327
-
[29]
C. Hofer, R. Kwitt, M. Niethammer, Y. Höller, E. Trinka, A. Uhl, Constructing Shape Spaces from a Topological Perspective, Springer International Publishing, 2017, p. 106–118.doi:10.1007/ 978-3-319-59050-9_9. URLhttp://dx.doi.org/10.1007/978-3-319-59050-9_9
-
[30]
119–128.doi:10.1007/978-3-030-87444-5_12
A.Saadat-Yazdi, R.Andreeva, R.Sarkar, Topologicaldetectionofalzheimer’sdiseaseusingbetticurves, in: Interpretability of Machine Intelligence in Medical Image Computing and Topological Data Analysis andItsApplicationsforMedicalData(TDA4MedicalData)WorkshopatMICCAI,Vol.12929ofLecture Notes in Computer Science, Springer, 2021, pp. 119–128.doi:10.1007/978-3-0...
-
[31]
S. C. Brüningk, F. Hensel, L. P. Lukas, M. Kuijs, C. R. Jutzeler, B. Rieck, Back to the basics with inclusion of clinical domain knowledge - a simple, scalable and effective model of alzheimer’s disease classification, in: K. Jung, S. Yeung, M. Sendak, M. Sjoding, R. Ranganath (Eds.), Proceedings of the 6th Machine Learning for Healthcare Conference, Vol....
work page 2021
-
[32]
T. Glozman, J. Solomon, F. Pestilli, L. Guibas, Shape-attributes of brain structures as biomarkers for alzheimer’s disease, Journal of Alzheimer’s Disease 56 (1) (2016) 287–295.doi:10.3233/jad-160900. URLhttp://dx.doi.org/10.3233/JAD-160900
-
[33]
Y. Singh, E. Quaia, Unraveling the invisible: Topological data analysis as the new frontier in radiology’s diagnostic arsenal, Tomography 11 (1) (2025) 6.doi:10.3390/tomography11010006. URLhttp://dx.doi.org/10.3390/tomography11010006
-
[34]
Bubenik, Statistical topological data analysis using persistence landscapes 16 (2015) 77–102
P. Bubenik, Statistical topological data analysis using persistence landscapes 16 (2015) 77–102. URLhttps://www.jmlr.org/papers/v16/bubenik15a.html
work page 2015
-
[35]
H. Edelsbrunner, E. P. Mücke, Three-dimensional alpha shapes 13 (1) (1994) 43–72.doi:10.1145/ 174462.156635. URLhttps://dl.acm.org/doi/10.1145/174462.156635 19
-
[36]
The GUDHI library: Simplicial complexes and persistent homology
C. Maria, J.-D. Boissonnat, et al., The GUDHI library: Simplicial complexes and persistent homology, in: International Congress on Mathematical Software (ICMS), Vol. 8592 of Lecture Notes in Computer Science, Springer, 2014, pp. 167–174.doi:10.1007/978-3-662-44199-2_28. URLhttps://link.springer.com/chapter/10.1007/978-3-662-44199-2_28
-
[37]
T. G. Project, GUDHI User and Reference Manual, 3rd Edition, GUDHI Editorial Board, 2025. URLhttps://gudhi.inria.fr/doc/3.11.0/
work page 2025
-
[38]
O. Esteban, C. J. Markiewicz, R. Blair, R. A. Poldrack, K. J. Gorgolewski, smriprep: Structural mri preprocessing workflows (Dec. 2024).doi:10.5281/zenodo.14532379. URLhttps://doi.org/10.5281/zenodo.14532379
-
[39]
C. R. Jack, M. A. Bernstein, N. C. Fox, P. Thompson, G. Alexander, D. Harvey, B. Borowski, P. J. Britson, J. L. Whitwell, C. Ward, A. M. Dale, J. P. Felmlee, J. L. Gunter, D. L. Hill, R. Killiany, N. Schuff, S. Fox-Bosetti, C. Lin, C. Studholme, C. S. DeCarli, G. Krueger, H. A. Ward, G. J. Metzger, K. T. Scott, R. Mallozzi, D. Blezek, J. Levy, J. P. Debbi...
-
[40]
Y. Singh, C. Farrelly, Q. A. Hathaway, G. Carlsson, Persistence barcodes: A novel approach to reducing bias in radiological analysis, Oncotarget 15 (1) (2024) 784–786.doi:10.18632/oncotarget.28667. URLhttp://dx.doi.org/10.18632/oncotarget.28667
-
[41]
Edelsbrunner, Alpha shapes — a survey, 2009
H. Edelsbrunner, Alpha shapes — a survey, 2009. URLhttps://api.semanticscholar.org/CorpusID:13439274
work page 2009
-
[42]
H. Braak, E. Braak, Staging of alzheimer’s disease-related neurofibrillary changes, Neurobiology of Aging 16 (3) (1995) 271–278.doi:10.1016/0197-4580(95)00021-6. URLhttp://dx.doi.org/10.1016/0197-4580(95)00021-6
-
[43]
B. C. Dickerson, A. Bakkour, D. H. Salat, E. Feczko, J. Pacheco, D. N. Greve, F. Grodstein, C. I. Wright, D. Blacker, H. D. Rosas, R. A. Sperling, A. Atri, J. H. Growdon, B. T. Hyman, J. C. Morris, B. Fischl, R. L. Buckner, The cortical signature of alzheimer’s disease: Regionally specific cortical thinningrelatestosymptomseverityinverymildtomildaddementi...
-
[44]
T. R. Stoub, L. deToledo Morrell, B. C. Dickerson, Parahippocampal white matter volume predicts alzheimer’s disease risk in cognitively normal old adults, Neurobiology of Aging 35 (8) (2014) 1855–1861. doi:10.1016/j.neurobiolaging.2014.01.153. URLhttp://dx.doi.org/10.1016/j.neurobiolaging.2014.01.153
-
[45]
F. Chazal, V. de Silva, S. Oudot, Persistence stability for geometric complexes, Geometriae Dedicata 173 (1) (2013) 193–214.doi:10.1007/s10711-013-9937-z. URLhttp://dx.doi.org/10.1007/s10711-013-9937-z
-
[46]
P. S. Aisen, R. C. Petersen, M. C. Donohue, A. Gamst, R. Raman, R. G. Thomas, S. Walter, J. Q. Trojanowski, L. M. Shaw, L. A. Beckett, C. R. Jack, W. Jagust, A. W. Toga, A. J. Saykin, J. C. Morris, R. C. Green, M. W. Weiner, Clinical core of the alzheimer’s disease neuroimaging initiative: Progress and plans, Alzheimer’s & Dementia 6 (3) (2010) 239–246.do...
-
[47]
Supplementary Material In this section we provide complementary analyses for our pipelines. We discuss correlations of our features with clinical measures to better understand the possible clinical use cases of our framework. We then provide a stratification analysis for Pipeline 1; this is needed to assess confounding effects with well-known covariatesli...
-
[48]
MeanL 1 curves(±SD)acrossanatomicalaxestoassesswhetherdisease-relatedmorphologicalpatterns persist after controlling for confounds
-
[49]
Effect sizes (Cohen’sd) on totalL1-curve AUC (summed across axes) to quantify the magnitude of group differences within each stratum. 5.2.2. Results Differences inL1 curves between CN and AD subjects persisted after stratification by both age and TIV (Figures 13, 14). Within every stratum, AD subjects showed lowerL1-curve AUC than CN across all three anat...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.