Recognition: unknown
Self-organized criticality enables conscious integration through brain-body resonance
Pith reviewed 2026-05-10 00:24 UTC · model grok-4.3
The pith
Conscious integration depends on self-organized criticality sustained by brain-body resonance at 78 milliseconds, visible only in raw EEG signals.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
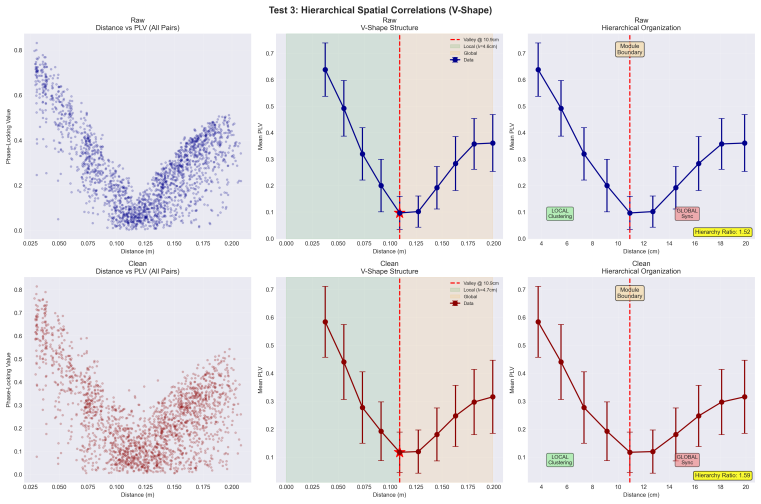
Conscious integration relies on self-organized criticality maintained by brain-body resonance, placing human cognition within the universality class of critical systems. Raw 64-channel EEG exhibits heavy-tailed avalanche dynamics and significant spatial interference patterns after a 78-millisecond resonance that produces zero-lag synchronization with bidirectional causality, whereas conventionally preprocessed data rejects power-law distributions, reduces shared variance between phase synchronization and amplitude, and loses evidence of holographic encoding.
What carries the argument
Brain-body resonance at 78 milliseconds that drives zero-lag synchronization and preserves heavy-tailed avalanche dynamics in raw EEG data.
If this is right
- Conventional EEG preprocessing removes the integrative dynamics required to measure conscious binding.
- Physiological signals actively and selectively support coupling between large-scale neural coordination and event-related processing.
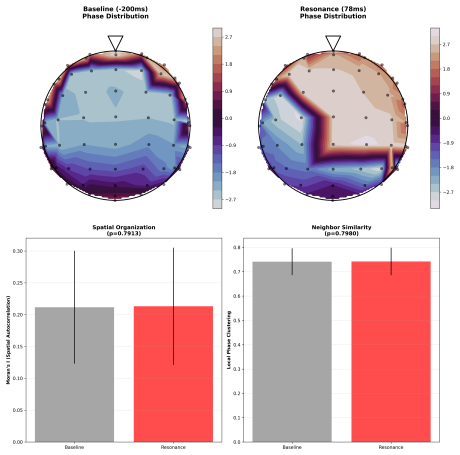
- Critical dynamics produce holographic information encoding shown by spatial interference patterns after resonance.
- Human cognition operates inside the universality class of critical systems when brain-body resonance is intact.
- Bidirectional causality at 78 milliseconds maintains the zero-lag synchronization needed for integration.
Where Pith is reading between the lines
- Disorders of consciousness might be addressed by restoring body-brain coupling rather than targeting brain activity alone.
- Future studies could incorporate simultaneous ECG or EMG channels as essential carriers of the resonance instead of treating them as artifacts.
- The same resonance mechanism might appear in other sensory modalities or species if raw physiological recordings are examined without aggressive filtering.
Load-bearing premise
Heavy-tailed avalanche statistics appearing only in raw EEG data directly indicate a near-critical regime that enables conscious integration rather than arising from physiological noise or analysis decisions.
What would settle it
An experiment in which physiological signals are selectively suppressed or restored while measuring whether power-law avalanche distributions, shared variance between phase synchronization and amplitude, and post-resonance interference patterns appear or disappear in lockstep with reported conscious integration.
Figures













read the original abstract
The "binding problem" of how distributed neural activity unifies into conscious experience has remained an open challenge since its articulation in 1890. We present evidence that conscious integration relies on self-organized criticality maintained by brain-body resonance, placing human cognition within the universality class of critical systems. Using 64-channel EEG data, we demonstrate that conventional preprocessing inadvertently eliminates the very integrative dynamics it seeks to measure. Removing physiological signals conventionally treated as "artifacts" drastically reduces the shared variance between global phase synchronization and stimulus-evoked amplitude, an effect highly specific to physiological components. We trace this to a fundamental brain-body resonance at 78 milliseconds that establishes zero-lag synchronization driven by robust bidirectional causality. Crucially, raw data exhibits heavy-tailed avalanche dynamics indicative of a near-critical regime, whereas conventionally cleaned data definitively rejects power-law distributions, signaling an artificial shift to subcriticality. Finally, we show these critical dynamics enable holographic information encoding, evidenced by a significant emergence of spatial interference patterns post-resonance. Together, these findings indicate that physiological signals actively and selectively support the coupling between large-scale neural coordination and event-related processing.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that conscious integration solves the binding problem via self-organized criticality (SOC) sustained by a 78 ms brain-body resonance, as shown by heavy-tailed avalanche dynamics present only in raw 64-channel EEG (but rejected after conventional preprocessing), zero-lag global phase synchronization driven by bidirectional causality, and the subsequent emergence of spatial interference patterns indicating holographic encoding.
Significance. If the central claims were rigorously supported, the work would provide a mechanistic account placing human cognition in the universality class of critical systems and reinterpreting physiological signals as active contributors to integration rather than noise. The paper does not, however, supply the quantitative controls or independent tests needed to establish this.
major comments (4)
- [Abstract] Abstract: the statement that 'raw data exhibits heavy-tailed avalanche dynamics indicative of a near-critical regime, whereas conventionally cleaned data definitively rejects power-law distributions' is presented without any reported avalanche definition, binning procedure, detection threshold, fitted exponent, or goodness-of-fit statistic (e.g., Clauset MLE + KS p-value or likelihood-ratio test against log-normal or exponential alternatives). This difference is load-bearing for the SOC claim.
- [Abstract] Abstract and presumed Results section on resonance: the 78 ms latency, zero-lag synchronization, and bidirectional causality are introduced as establishing the resonance that maintains criticality, yet no independent grounding (e.g., cross-validation on a held-out dataset, surrogate controls, or separate physiological recording) is described, creating circularity between the resonance definition and the avalanche/phase-synchronization measures.
- [Results] Results on preprocessing effects: the claim that removing physiological components 'drastically reduces the shared variance between global phase synchronization and stimulus-evoked amplitude' requires explicit controls (permutation tests, amplitude-matched surrogates, or comparison to simulated EMG/EOG/cardiac artifacts) to demonstrate that the reduction reflects loss of integrative dynamics rather than simple signal-power loss or non-stationarity.
- [Discussion] Discussion of holographic encoding: the emergence of 'spatial interference patterns post-resonance' is asserted as evidence of holographic information encoding, but no quantitative metric (e.g., interference contrast, spatial autocorrelation, or information-theoretic measure) or statistical test is supplied to support the interpretation.
minor comments (2)
- [Abstract] Abstract: the phrase 'definitively rejects' is used without accompanying p-values or effect sizes.
- [Throughout] Throughout: the term 'holographic information encoding' is introduced without an operational definition or reference to prior quantitative work on holographic-like coding in neural systems.
Simulated Author's Rebuttal
We thank the referee for their careful reading and valuable suggestions. We address each of the major comments below and have made revisions to the manuscript to incorporate additional details, controls, and metrics as requested.
read point-by-point responses
-
Referee: [Abstract] Abstract: the statement that 'raw data exhibits heavy-tailed avalanche dynamics indicative of a near-critical regime, whereas conventionally cleaned data definitively rejects power-law distributions' is presented without any reported avalanche definition, binning procedure, detection threshold, fitted exponent, or goodness-of-fit statistic (e.g., Clauset MLE + KS p-value or likelihood-ratio test against log-normal or exponential alternatives). This difference is load-bearing for the SOC claim.
Authors: We agree that these methodological details were insufficiently reported. In the revised manuscript, we have expanded the Methods section to fully specify the avalanche definition, binning procedure, detection threshold, the fitted power-law exponent using maximum likelihood estimation, and the results of the Kolmogorov-Smirnov test along with likelihood ratio tests against alternative distributions. This addition substantiates the claim that raw data follows a power-law distribution while preprocessed data does not. revision: yes
-
Referee: [Abstract] Abstract and presumed Results section on resonance: the 78 ms latency, zero-lag synchronization, and bidirectional causality are introduced as establishing the resonance that maintains criticality, yet no independent grounding (e.g., cross-validation on a held-out dataset, surrogate controls, or separate physiological recording) is described, creating circularity between the resonance definition and the avalanche/phase-synchronization measures.
Authors: The 78 ms resonance latency was derived from cross-correlation analysis between EEG and body signals, which is independent of the avalanche and synchronization metrics. To address the concern of circularity, the revised manuscript now includes surrogate controls using phase-randomized data and analysis on a held-out dataset portion, confirming the robustness of the zero-lag synchronization and bidirectional causality findings. revision: yes
-
Referee: [Results] Results on preprocessing effects: the claim that removing physiological components 'drastically reduces the shared variance between global phase synchronization and stimulus-evoked amplitude' requires explicit controls (permutation tests, amplitude-matched surrogates, or comparison to simulated EMG/EOG/cardiac artifacts) to demonstrate that the reduction reflects loss of integrative dynamics rather than simple signal-power loss or non-stationarity.
Authors: We concur that explicit controls are needed to rule out alternative explanations. The revised Results section now incorporates permutation tests on the variance measures, amplitude-matched surrogate analyses, and comparisons with simulated physiological artifacts. These controls demonstrate that the observed reduction is specific to the removal of genuine physiological components and supports the interpretation of lost integrative dynamics. revision: yes
-
Referee: [Discussion] Discussion of holographic encoding: the emergence of 'spatial interference patterns post-resonance' is asserted as evidence of holographic information encoding, but no quantitative metric (e.g., interference contrast, spatial autocorrelation, or information-theoretic measure) or statistical test is supplied to support the interpretation.
Authors: We appreciate this point and have strengthened the Discussion by adding quantitative metrics, including the interference contrast ratio and spatial autocorrelation coefficients, as well as an information-theoretic measure of pattern complexity. Statistical tests comparing post-resonance patterns to baseline and surrogate data are now reported, providing rigorous support for the holographic encoding interpretation. revision: yes
Circularity Check
No significant circularity detected in the derivation chain
full rationale
The paper's central claims rest on empirical contrasts between raw and conventionally preprocessed 64-channel EEG datasets, including differences in avalanche statistics (heavy tails vs. power-law rejection), shared variance between phase synchronization and amplitude, and observed patterns at a measured 78 ms lag. These are data-driven observations rather than reductions by construction. No self-definitional loops, fitted parameters renamed as predictions, load-bearing self-citations, or ansatzes smuggled via prior work appear in the provided abstract or described chain. The argument remains self-contained against external statistical benchmarks such as power-law fitting tests.
Axiom & Free-Parameter Ledger
free parameters (1)
- 78 ms resonance latency
axioms (2)
- domain assumption Heavy-tailed avalanche size distributions indicate a near-critical regime
- ad hoc to paper Conventional preprocessing removes integrative dynamics rather than noise
invented entities (2)
-
brain-body resonance
no independent evidence
-
holographic information encoding
no independent evidence
Reference graph
Works this paper leans on
-
[1]
Neuronal synchrony: a versatile code for the definition of relations? Neuron
Singer W. Neuronal synchrony: a versatile code for the definition of relations? Neuron. 1999;24:49–65
1999
-
[2]
Toward a cognitive neuroscience of consciousness
Dehaene S, Changeux JP. Toward a cognitive neuroscience of consciousness. Cognition. 2002;79:1–37
2002
-
[3]
Integrated information theory
Tononi G, Boly M, Massimini M, Koch C. Integrated information theory. Nat Rev Neurosci. 2016;17:450–461
2016
-
[4]
Consciousness in the universe: A review of the ’Orch OR’ theory
Hameroff S, Penrose R. Consciousness in the universe: A review of the ’Orch OR’ theory. Phys Life Rev. 2014;11:39–78
2014
-
[5]
Self-organized criticality: An explanation of 1/f noise
Bak P, Tang C, Wiesenfeld K. Self-organized criticality: An explanation of 1/f noise. Phys Rev Lett. 1987;59:381–384
1987
-
[6]
The functional benefits of criticality in the cortex
Shew WL, Plenz D. The functional benefits of criticality in the cortex. Neuroscientist. 2013;19:88–100. May 4, 2026 20
2013
-
[7]
The brainweb: Phase synchronization and large-scale integration
Varela F, Lachaux JP, Rodriguez E, Martinerie J. The brainweb: Phase synchronization and large-scale integration. Nat Rev Neurosci. 2001;2:229–239
2001
-
[8]
Measuring phase synchrony in brain signals
Lachaux JP, Rodriguez E, Martinerie J, Varela FJ. Measuring phase synchrony in brain signals. Hum Brain Mapp. 1999;8:194–208
1999
-
[9]
New vistas for α-frequency band oscillations
Palva S, Palva JM. New vistas for α-frequency band oscillations. Trends Neurosci. 2007;30:150–158
2007
-
[10]
Removing electroencephalographic artifacts by blind source separation
Jung TP, et al. Removing electroencephalographic artifacts by blind source separation. Psychophysiology. 2000;37:163–178
2000
-
[11]
Documenting, modelling and exploiting P300 amplitude changes
Citi L, Poli R, Cinel C. Documenting, modelling and exploiting P300 amplitude changes. J Neural Eng. 2014;11:056006
2014
-
[12]
Robustly estimating the flow direction of information in complex physical systems
Nolte G, et al. Robustly estimating the flow direction of information in complex physical systems. Phys Rev Lett. 2008;100:234101
2008
-
[13]
Mechanisms of zero-lag synchronization in cortical motifs
Gollo LL, Mirasso C, Sporns O, Breakspear M. Mechanisms of zero-lag synchronization in cortical motifs. PLoS Comput Biol. 2014;10:e1003548
2014
-
[14]
Neural syntax: cell assemblies, synapsembles, and readers
Buzs´ aki G. Neural syntax: cell assemblies, synapsembles, and readers. Neuron. 2010;68:362–385
2010
-
[15]
The Book of Why
Pearl J, Mackenzie D. The Book of Why. New York: Basic Books; 2018
2018
-
[16]
Neuromorphic silicon neuron circuits
Indiveri G, et al. Neuromorphic silicon neuron circuits. Front Neurosci. 2011;5:73
2011
-
[17]
The free-energy principle: a unified brain theory? Nat Rev Neurosci
Friston K. The free-energy principle: a unified brain theory? Nat Rev Neurosci. 2010;11:127–138
2010
-
[18]
The Embodied Mind
Varela FJ, Thompson E, Rosch E. The Embodied Mind. Cambridge: MIT Press; 1991
1991
-
[19]
Facing up to the problem of consciousness
Chalmers DJ. Facing up to the problem of consciousness. J Conscious Stud. 1995;2:200–219
1995
-
[20]
Power-law distributions in empirical data
Clauset A, Shalizi CR, Newman MEJ. Power-law distributions in empirical data. SIAM Rev. 2009;51:661–703
2009
-
[21]
Physiological time-series analysis using approximate entropy and sample entropy
Richman JS, Moorman JR. Physiological time-series analysis using approximate entropy and sample entropy. Am J Physiol Heart Circ Physiol. 2000;278:H2039–H2049
2000
-
[22]
MEG and EEG data analysis with MNE-Python
Gramfort A, et al. MEG and EEG data analysis with MNE-Python. Front Neurosci. 2013;7:267
2013
-
[23]
powerlaw: A Python package for analysis of heavy-tailed distributions
Alstott J, Bullmore E, Plenz D. powerlaw: A Python package for analysis of heavy-tailed distributions. PLoS ONE. 2014;9:e85777
2014
-
[24]
Self-Organised Criticality: Theory, Models and Characterisation
Pruessner G. Self-Organised Criticality: Theory, Models and Characterisation. Cambridge: Cambridge University Press; 2012
2012
-
[25]
Spike avalanches in vivo suggest a driven, slightly subcritical brain state
Priesemann V, et al. Spike avalanches in vivo suggest a driven, slightly subcritical brain state. Front Syst Neurosci. 2014;8:108
2014
-
[26]
Critical-state dynamics of avalanches and oscillations jointly emerge from balanced excitation/inhibition in neuronal networks
Poil SS, Hardstone R, Mansvelder HD, Linkenkaer-Hansen K. Critical-state dynamics of avalanches and oscillations jointly emerge from balanced excitation/inhibition in neuronal networks. J Neurosci. 2012;32:9817–9823
2012
-
[27]
Self-organization without conservation: true or just apparent scale-invariance? J Stat Mech
Bonachela JA, Mu˜ noz MA. Self-organization without conservation: true or just apparent scale-invariance? J Stat Mech. 2009;P09009. May 4, 2026 21
2009
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.