A trick of the tail: how electrostatics helps a DNA repair enzyme to localize on nucleosomes
Pith reviewed 2026-06-29 00:43 UTC · model grok-4.3
The pith
An arginine anchor motif on the N-terminal tail of UDG favors its electrostatic binding to acidic patches on nucleosomes.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The presence of an arginine anchor motif on the N-terminal tail of UDG can favor its localization on nucleosomes by binding to their acidic patches on their top and bottom surfaces via electrostatic interactions. We argue that this mechanism can play a key role in the detection of uracil defects in nucleosomal DNA.
What carries the argument
The arginine anchor motif on the N-terminal tail of UDG, which performs electrostatic binding to nucleosome acidic patches to promote localization.
If this is right
- UDG can reach nucleosomal DNA via surface binding on top and bottom faces before scanning for uracil.
- The electrostatic mechanism supports base-excision repair initiation inside chromatin without requiring immediate DNA unwrapping.
- Other base-excision repair enzymes may use analogous tail motifs for nucleosome targeting.
- Uracil detection efficiency in condensed chromatin depends in part on this initial electrostatic docking step.
Where Pith is reading between the lines
- Disrupting the arginine motif in living cells would be expected to slow overall uracil repair rates if the localization step is rate-limiting.
- The same acidic-patch binding could be tested for competition with other chromatin proteins that recognize the same patches.
- Simulations of the full nucleosome plus UDG tail could quantify the binding energy contribution relative to thermal fluctuations.
Load-bearing premise
Electrostatic attraction from the arginine motif is sufficient to favor UDG localization on nucleosomes instead of being outweighed by other forces or steric constraints.
What would settle it
A direct measurement showing that mutating the arginine residues in the motif produces no measurable change in UDG binding probability or residence time on nucleosomes would refute the proposed localization advantage.
Figures

read the original abstract
Electrostatic interactions are key to the recognition processes of proteins and DNA and have been previously documented for the action of repair enzymes. Uracil-DNA glycosylase (UDG) is the first in a sequence of enzymes that act in the base-excision repair process (BER) and whose task is the extraction of uracil bases from nuclear DNA. The question of how the molecule targets uracil bases in chromatin, in particular in the condensed protein-DNA complexes of nucleosomes, has only recently become a subject of detailed studies. Here we show that the presence of an arginine anchor motif on the N-terminal tail of UDG can favor its localization on nucleosomes by binding to their acidic patches on their top and bottom surfaces via electrostatic interactions. We argue that this mechanism can play a key role in the detection of uracil defects in nucleosomal DNA.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that an arginine anchor motif on the N-terminal tail of uracil-DNA glycosylase (UDG) can favor localization on nucleosomes by electrostatic binding to acidic patches on the top and bottom surfaces, and argues this mechanism can play a key role in detecting uracil defects in nucleosomal DNA during base-excision repair.
Significance. If the electrostatic localization mechanism holds, the work would add to understanding of how BER enzymes access damage sites within chromatin by leveraging known acidic-patch recognition motifs. The deliberately modest phrasing ('can favor', 'can play a key role') avoids overclaiming dominance over steric or other factors.
major comments (1)
- [Abstract] Abstract: the central claim that the arginine anchor 'can favor' localization and 'can play a key role' is presented without any quantitative threshold, binding-energy estimate, simulation protocol, or comparison to competing interactions. This makes it impossible to evaluate whether the electrostatic contribution is load-bearing or merely permissive.
Simulated Author's Rebuttal
We thank the referee for their review. We address the single major comment below.
read point-by-point responses
-
Referee: [Abstract] Abstract: the central claim that the arginine anchor 'can favor' localization and 'can play a key role' is presented without any quantitative threshold, binding-energy estimate, simulation protocol, or comparison to competing interactions. This makes it impossible to evaluate whether the electrostatic contribution is load-bearing or merely permissive.
Authors: We agree that the abstract, in its current form, is purely qualitative and does not supply the requested quantitative context. The main text of the manuscript reports the molecular-dynamics protocol and the electrostatic binding-energy estimates obtained from those simulations. To allow readers to assess whether the interaction is load-bearing, we will revise the abstract to include a concise statement of the simulation approach and the magnitude of the computed electrostatic contribution relative to other factors. revision: yes
Circularity Check
No significant circularity in derivation chain
full rationale
The paper advances a qualitative biophysical argument that an arginine anchor motif enables electrostatic localization of UDG on nucleosome acidic patches. No equations, parameter fits presented as predictions, self-citation chains, or ansatzes that reduce the central claim to its own inputs appear in the abstract or described structure. The mechanism is offered as a plausible contribution consistent with known nucleosome features rather than a derived result forced by internal definitions or prior self-references.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
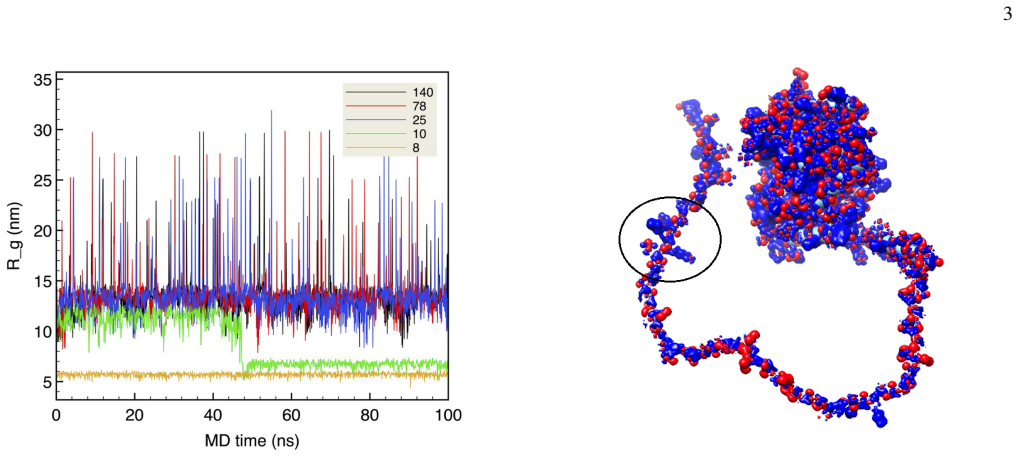
Conception of our study.Our study elucidates three aspects of the interaction between the full UDG molecule and the nucle- osome
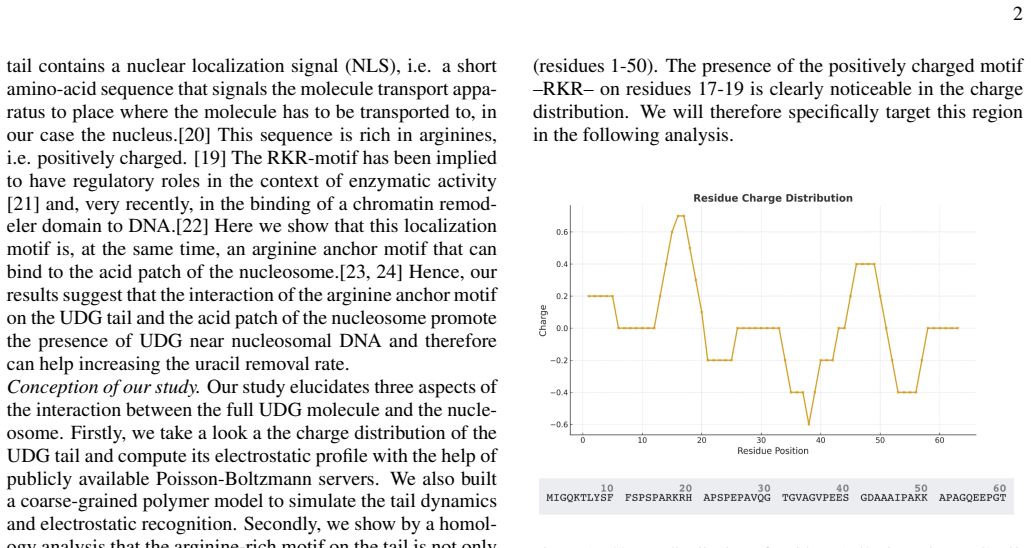
and, very recently, in the binding of a chromatin remod- eler domain to DNA.[22] Here we show that this localization motif is, at the same time, an arginine anchor motif that can bind to the acid patch of the nucleosome.[23, 24] Hence, our results suggest that the interaction of the arginine anchor motif on the UDG tail and the acid patch of the nucleosom...
-
[2]
sequence alignment similarity to the reference UDG motif
-
[3]
the presence of an RKR-like central motif
-
[4]
overall peptide charge
-
[5]
The score is constructed in detail as follows:
enrichment in serine and proline residues characteristic of intrinsically disordered regions. The score is constructed in detail as follows:
-
[6]
Alignment similarity The sequence similarity to the reference UDG motif was evaluated over a window of 15 aligned residues. For each aligned position, a score was assigned according to residue similarity according to: • identical residue: +2 • conservative substitution (R↔K, D↔E, S↔T): +1 • non-conservative substitution:−1 The alignment score is obtained ...
-
[7]
The central arginine motif Because acidic-patch binding motifs are characterized by the presence of a central arginine anchor, the peptide win- dow was examined for the occurrence of anRKR-type motif or closely related variants. Residues located in the central re- gion of the alignment window (positions 7–11) were therefore analyzed according to the follo...
-
[8]
Charge contribution Electrostatic complementarity is a major contributor to acidic-patch recognition. The net charge of the peptide win- dow was therefore included as Scorecharge = (#Arg+#Lys)−(#Asp+#Glu) 15 (2) where the numerator corresponds to the net charge of the 15- residue window. As a high positive charge alone is not suf- ficient to guarantee aci...
-
[9]
These residues pro- mote conformational flexibility and allow the peptide to adapt its structure upon binding
Serine and proline enrichment Many acidic-patch interacting peptides originate from in- trinsically disordered regions, which frequently exhibit en- richment in serine and proline residues. These residues pro- mote conformational flexibility and allow the peptide to adapt its structure upon binding. This contribution was quantified as ScoreSerPro = (#Ser+...
-
[10]
Final composite score The four contributions were combined into a weighted composite score: Scoreglobal =0.5Score align +0.3Score center (4) +0.1Score charge +0.1Score SerPro The weighting scheme prioritizes sequence similarity and the presence of the central arginine motif, while charge and disor- der propensity act as secondary modifiers. This scoring f...
-
[11]
Dyprosome
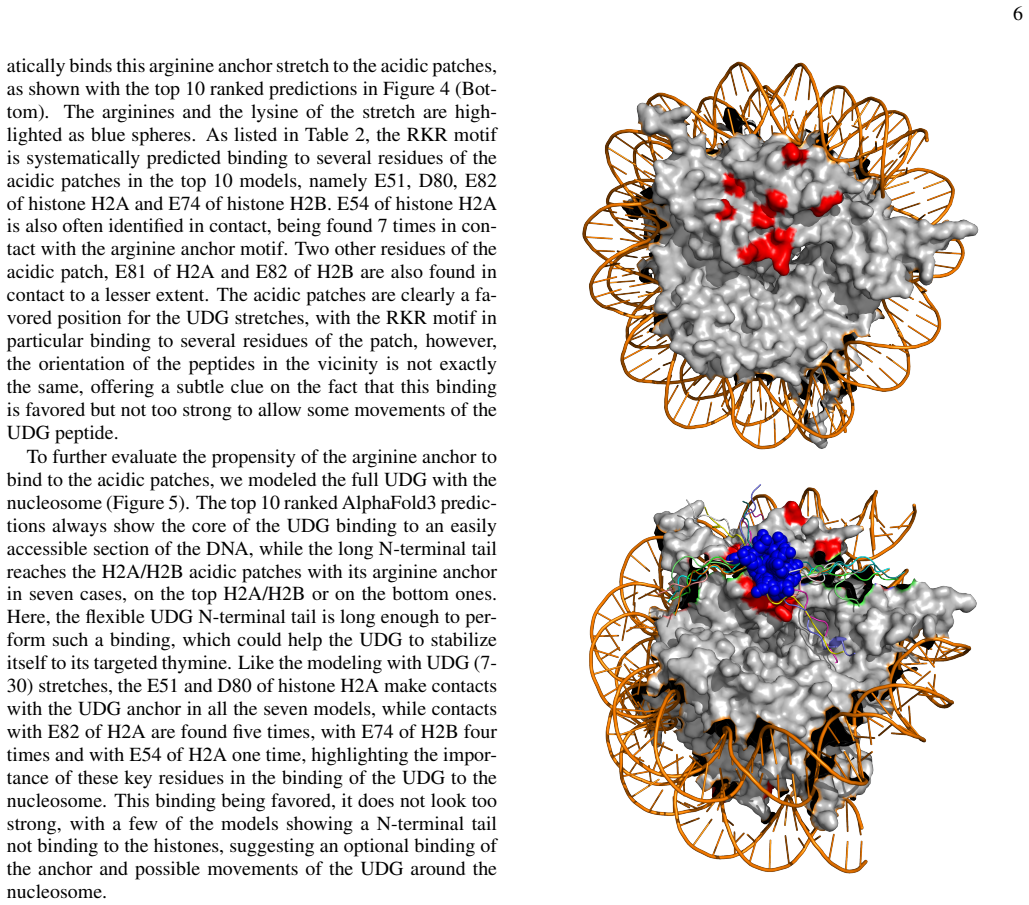
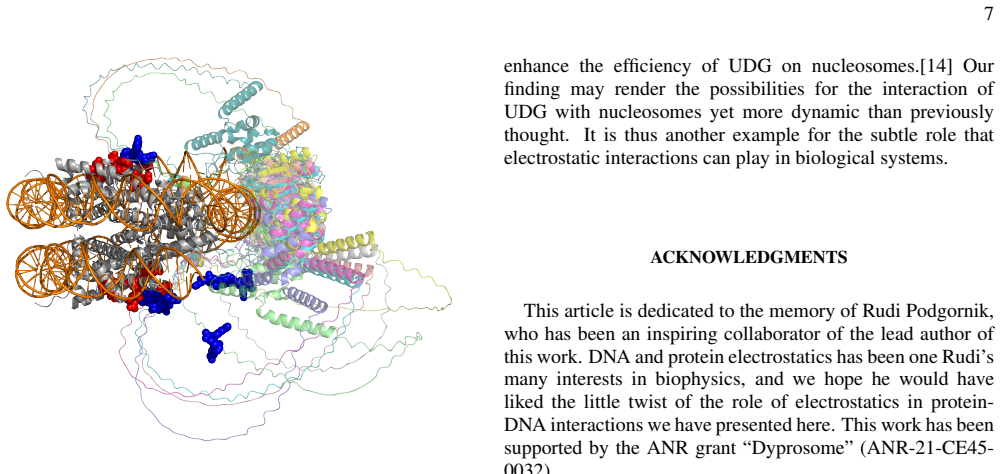
stretches, the E51 and D80 of histone H2A make contacts with the UDG anchor in all the seven models, while contacts with E82 of H2A are found five times, with E74 of H2B four times and with E54 of H2A one time, highlighting the impor- tance of these key residues in the binding of the UDG to the nucleosome. This binding being favored, it does not look too ...
-
[12]
H. E. Krokan and M. Bjørås, Cold Spring Harb Perspect Biol5, a012583 (2013)
2013
-
[13]
Slupphaug, C
G. Slupphaug, C. D. Mol, B. Kavli, A. S. Arvai, H. E. Krokan, and J. A. Tainer, Nature384, 87 (1996)
1996
-
[14]
Li and S
C. Li and S. Delaney, inDNA Repair, The Enzymes, V ol. 45, edited by L. Zhao and L. S. Kaguni (Academic Press, 2019) pp. 27–57
2019
-
[15]
M. Kong, E. C. Beckwitt, and B. Van Houten, DNA Repair (Amst)93, 102909 (2020)
2020
-
[16]
L. S. Bigman and Y . Levy, Annual Review of Biophysics52, 463 (2023)
2023
-
[17]
Saravanan, N
V . Saravanan, N. Raouraoua, G. Brysbaert, S. Giordano, M. F. Lensink, F. Cleri, and R. Blossey, Scientific Reports15, 7993 (2025)
2025
-
[18]
Cleri, S
F. Cleri, S. Giordano, and R. Blossey, J Mol Biol435, 168263 (2023)
2023
-
[19]
Dahirel, F
V . Dahirel, F. Paillusson, M. Jardat, M. Barbi, and J.-M. Victor, Phys. Rev. Lett.102, 228101 (2009)
2009
-
[20]
S. L. Cravens, M. Hobson, and J. T. Stivers, Biochemistry53, 7680 (2014)
2014
-
[21]
Y . Xie, C. B. Karki, J. Chen, D. Liu, and L. Li, Front Mol Biosci8, 718587 (2021)
2021
-
[22]
L. Li, C. Li, S. Sarkar, J. Zhang, S. Witham, Z. Zhang, L. Wang, N. Smith, M. Petukh, and E. Alexov, BMC Biophysics5, 9 (2012)
2012
-
[23]
Y . Ye, M. R. Stahley, J. Xu, J. I. Friedman, Y . Sun, J. N. McK- 8 night, J. J. Gray, G. D. Bowman, and J. T. Stivers, Biochemistry 51, 6028 (2012)
2012
-
[24]
D. J. Biechele-Speziale, T. B. Sutton, and S. Delaney, DNA Repair116, 103345 (2022)
2022
-
[25]
G. M. Barbosa and S. Delaney, DNA Repair150, 103852 (2025)
2025
-
[26]
S. Ghediri, P. A. P. Sarma, V . Saravanan, C. Ab- badie, R. Blossey, and F. Cleri, Biomolecules15(2025), 10.3390/biom15050649
-
[27]
S. Ghediri, R. Blossey, and F. Cleri, bioRxiv (2025), 10.1101/2025.04.17.649289
-
[28]
Vuzman, A
D. Vuzman, A. Azia, and Y . Levy, J Mol Biol396, 674 (2009)
2009
-
[29]
Rodriguez, B
G. Rodriguez, B. Orris, A. Majumdar, S. Bhat, and J. T. Stivers, DNA Repair (Amst)86, 102764 (2019)
2019
-
[30]
J. L. Perkins and L. Zhao, DNA Repair101, 103077 (2021)
2021
-
[31]
Otterlei, T
M. Otterlei, T. Haug, T. A. Nagelhus, G. Slupphaug, T. Lindmo, and H. E. Krokan, Nucleic Acids Research26, 4611 (1998)
1998
-
[32]
L. Wang, K. Zhang, L. Wu, S. Liu, H. Zhang, Q. Zhou, L. Tong, F. Sun, and Z. Fan, The Journal of Immunology188, 765 (2012)
2012
-
[33]
Yadav, P
Y . Yadav, P. Chyne, and B. Sudhamalla, Biochemical Journal 482, 1415 (2025)
2025
-
[34]
R. K. McGinty and S. Tan, Curr Opin Struct Biol71, 16 (2021)
2021
-
[35]
Zhang, J
H. Zhang, J. Eerland, V . Horn, R. Schellevis, and H. van Ingen, Scientific Reports11, 23013 (2021)
2021
-
[36]
Jumper, R
J. Jumper, R. Evans, A. Pritzel, T. Green, M. Figurnov, O. Ronneberger, K. Tunyasuvunakool, R. Bates, A. Žídek, A. Potapenko, A. Bridgland, C. Meyer, S. A. A. Kohl, A. J. Ballard, A. Cowie, B. Romera-Paredes, S. Nikolov, R. Jain, J. Adler, T. Back, S. Petersen, D. Reiman, E. Clancy, M. Zielin- ski, M. Steinegger, M. Pacholska, T. Berghammer, S. Boden- ste...
2021
-
[37]
P. Rice, I. Longden, and A. Bleasby, Trends Genet16, 276 (2000)
2000
-
[38]
Lei, P.-H
U. Lei, P.-H. Sun, and R. Pethig, Biomicrofluidics5, 044109 (2011)
2011
-
[39]
Salimi, K
E. Salimi, K. Braasch, M. Butler, D. J. Thomson, and G. E. Bridges, Biomicrofluidics10, 014111 (2016)
2016
-
[40]
Blossey,The Poisson-Boltzmann Equation: An Introduction (Springer, Cham, 2023)
R. Blossey,The Poisson-Boltzmann Equation: An Introduction (Springer, Cham, 2023)
2023
-
[41]
S. K. Panday, M. H. B. Shashikala, M. Koirala, S. Pahari, A. Chakvorty, Y . Peng, L. Li, Z. Jia, C. Li, and E. Alexov, Living Journal of Computational Molecular Science1, 10841 (2019)
2019
-
[42]
Jurrus, D
E. Jurrus, D. Engel, K. Star, K. Monson, J. Brandi, L. E. Fel- berg, D. H. Brookes, L. Wilson, J. Chen, K. Liles, M. Chun, P. Li, D. W. Gohara, T. Dolinsky, R. Konecny, D. R. Koes, J. E. Nielsen, T. Head-Gordon, W. Geng, R. Krasny, G.-W. Wei, M. J. Holst, J. A. McCammon, and N. A. Baker, Protein Sci- ence27, 112 (2018)
2018
-
[43]
P. D. Oleinikov, A. S. Fedulova, G. A. Armeev, N. A. Mo- torin, L. Singh-Palchevskaia, A. L. Sivkina, P. G. Feskin, G. S. Glukhov, D. A. Afonin, G. A. Komarova, M. P. Kirpichnikov, V . M. Studitsky, A. V . Feofanov, and A. K. Shaytan, Interna- tional Journal of Molecular Sciences24(2023)
2023
-
[44]
Abramson, J
J. Abramson, J. Adler, J. Dunger, R. Evans, T. Green, A. Pritzel, O. Ronneberger, L. Willmore, A. J. Ballard, J. Bambrick, S. W. Bodenstein, D. A. Evans, C.-C. Hung, M. O’Neill, D. Reiman, K. Tunyasuvunakool, Z. Wu, A. Žemgulyt˙e, E. Ar- vaniti, C. Beattie, O. Bertolli, A. Bridgland, A. Cherepanov, M. Congreve, A. I. Cowen-Rivers, A. Cowie, M. Figurnov, F...
2024
-
[45]
Raouraoua, C
N. Raouraoua, C. Mirabello, T. Véry, C. Blanchet, B. Wallner, M. F. Lensink, and G. Brysbaert, Nature Computational Sci- ence4, 824 (2024)
2024
-
[46]
Zhang and J
Y . Zhang and J. Skolnick, Proteins57, 702 (2004)
2004
-
[47]
R. Evans, M. O’Neill, A. Pritzel, N. Antropova, A. Se- nior, T. Green, A. Žídek, R. Bates, S. Blackwell, J. Yim, O. Ronneberger, S. Bodenstein, M. Zielinski, A. Bridg- land, A. Potapenko, A. Cowie, K. Tunyasuvunakool, R. Jain, E. Clancy, P. Kohli, J. Jumper, and D. Hassabis, bioRxiv (2022), 10.1101/2021.10.04.463034
-
[48]
The PyMOL molecular graphics system, version 1.8,
Schrödinger, LLC, “The PyMOL molecular graphics system, version 1.8,” (2015)
2015
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.