Morphogenesis driven by nematic defects in active biological networks
Pith reviewed 2026-06-28 07:46 UTC · model grok-4.3
The pith
Topological defects in nematic tissues control where protrusions form and growth occurs during morphogenesis.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
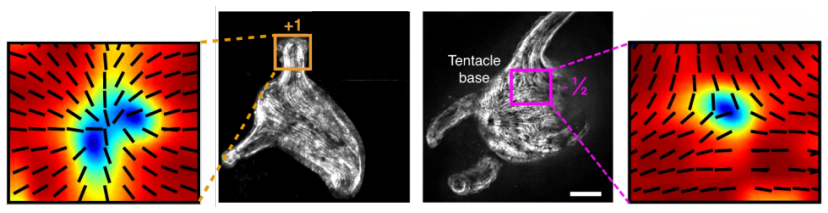
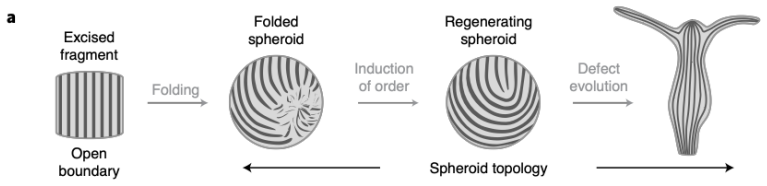
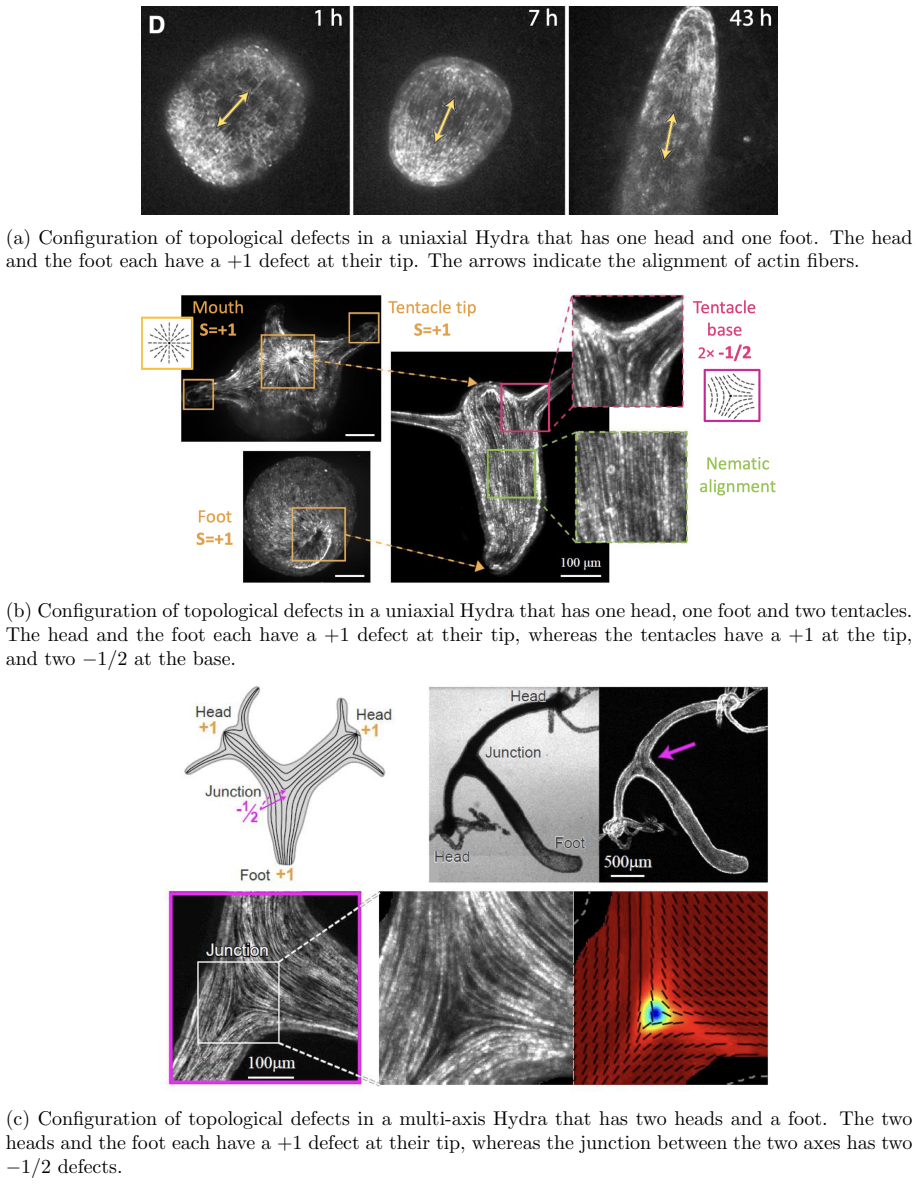
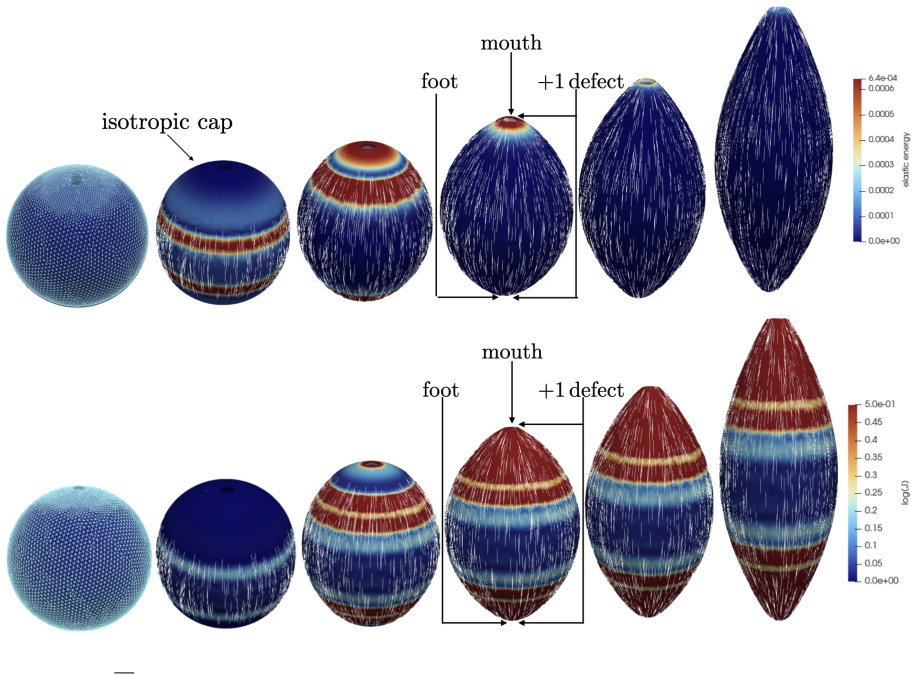
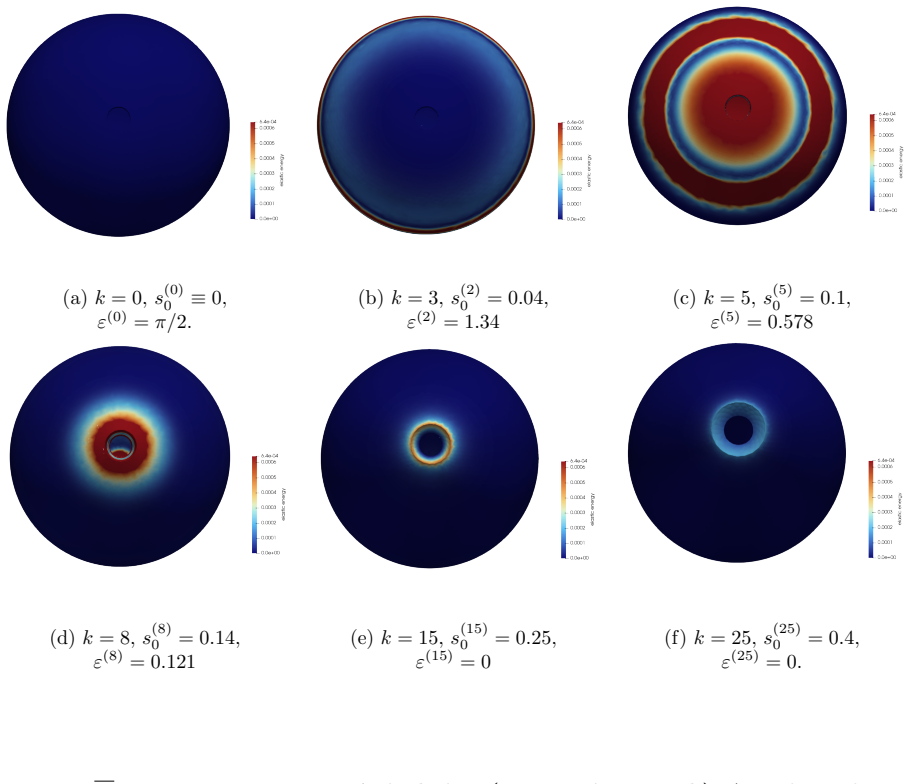
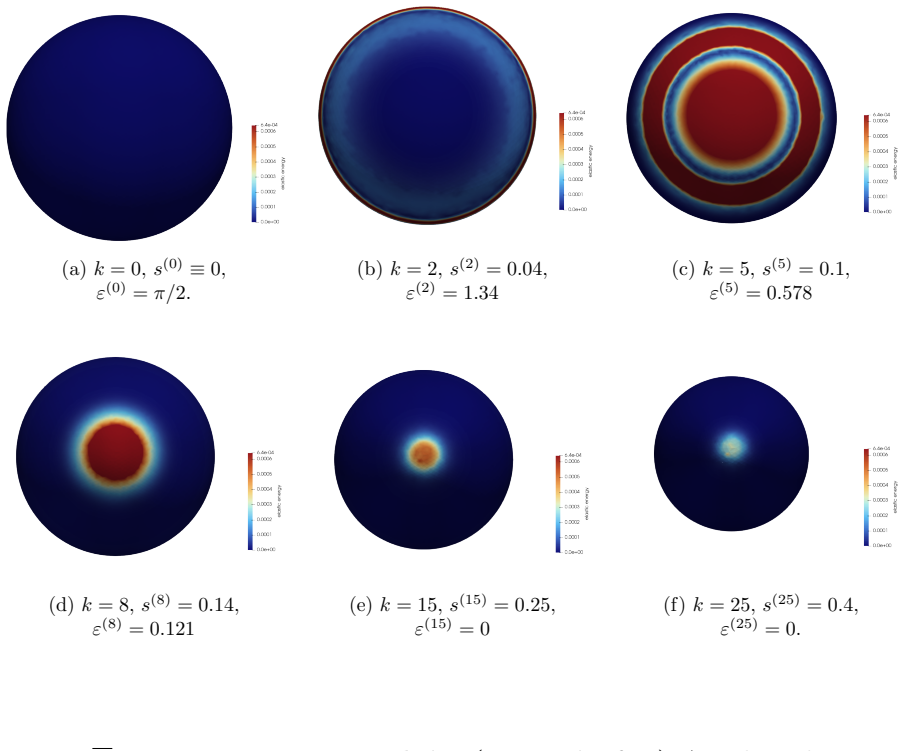
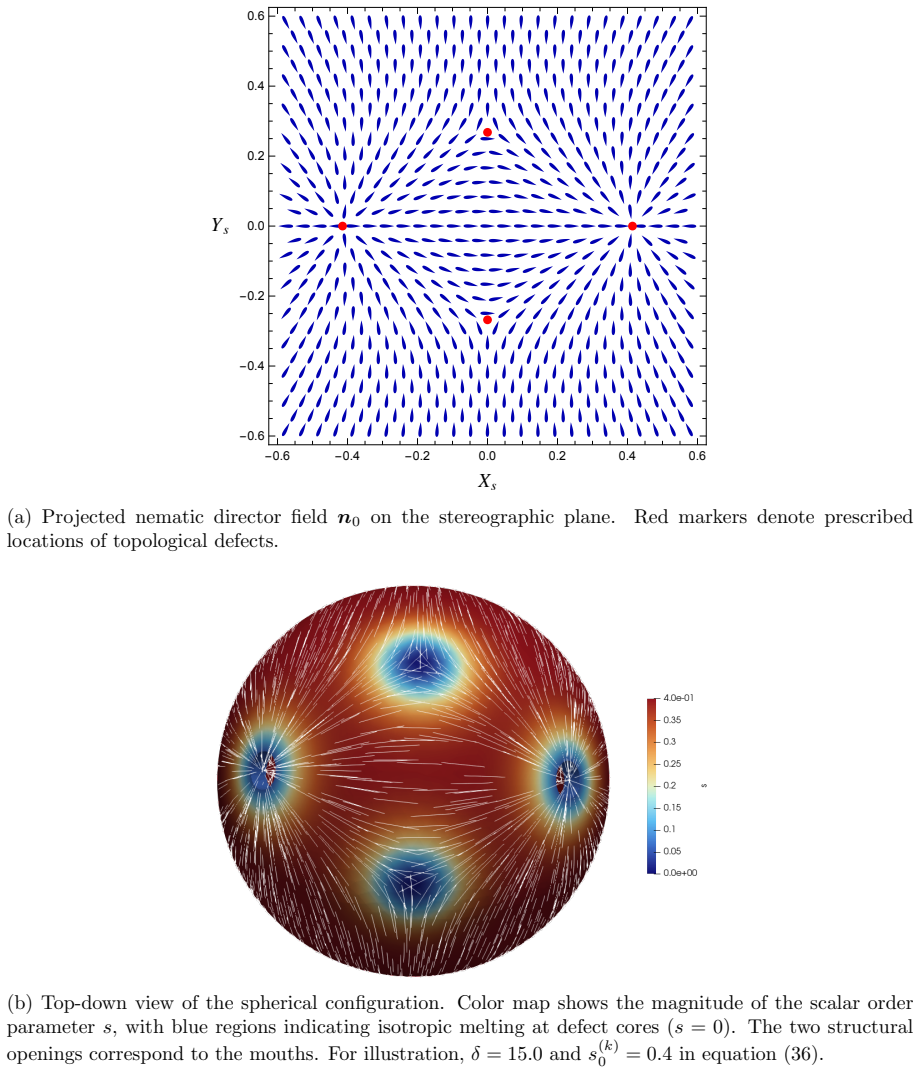
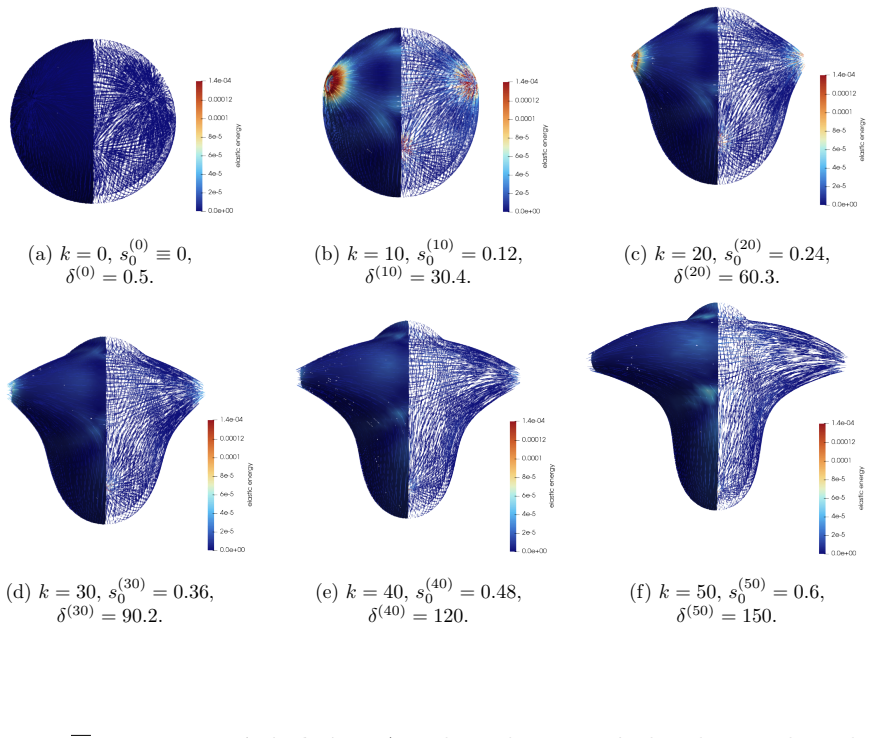
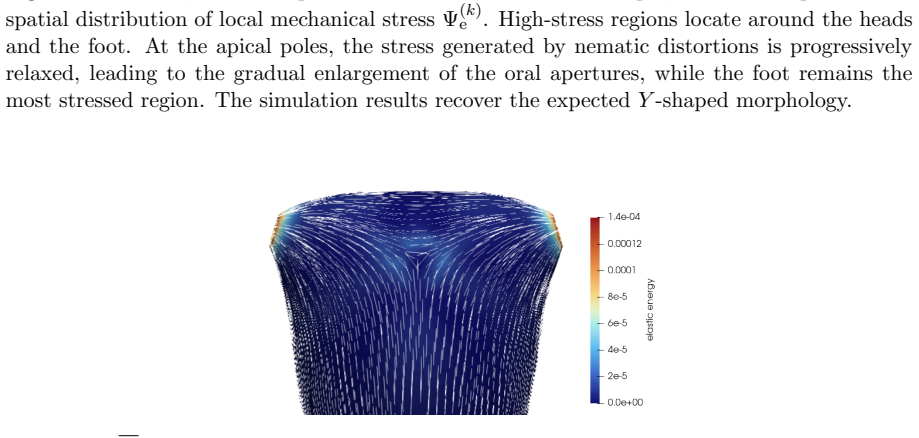
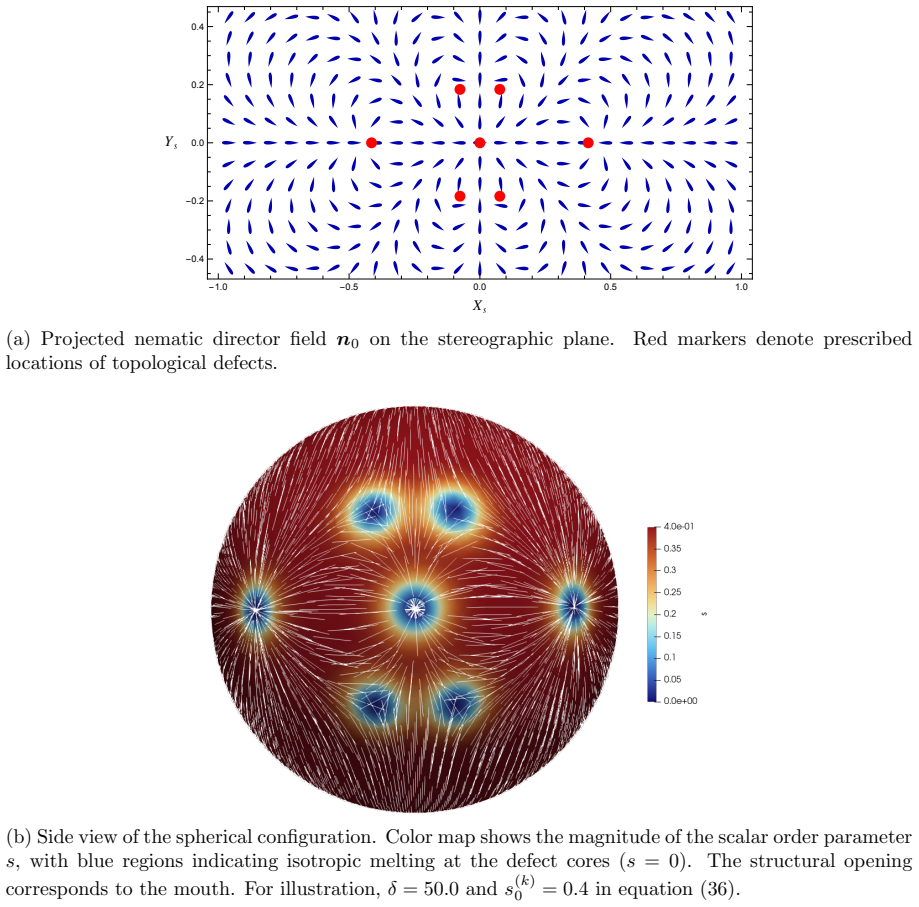
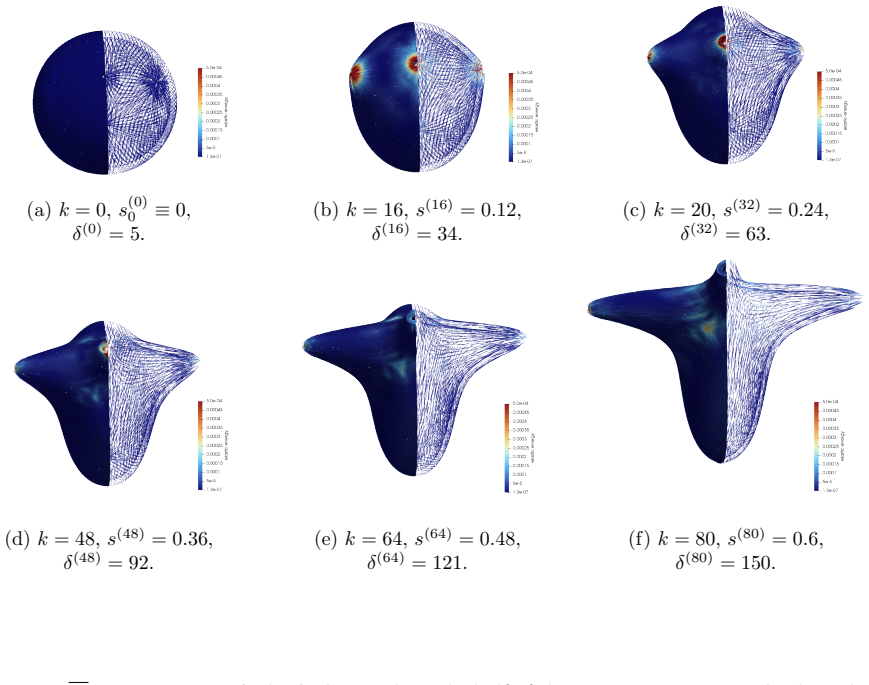
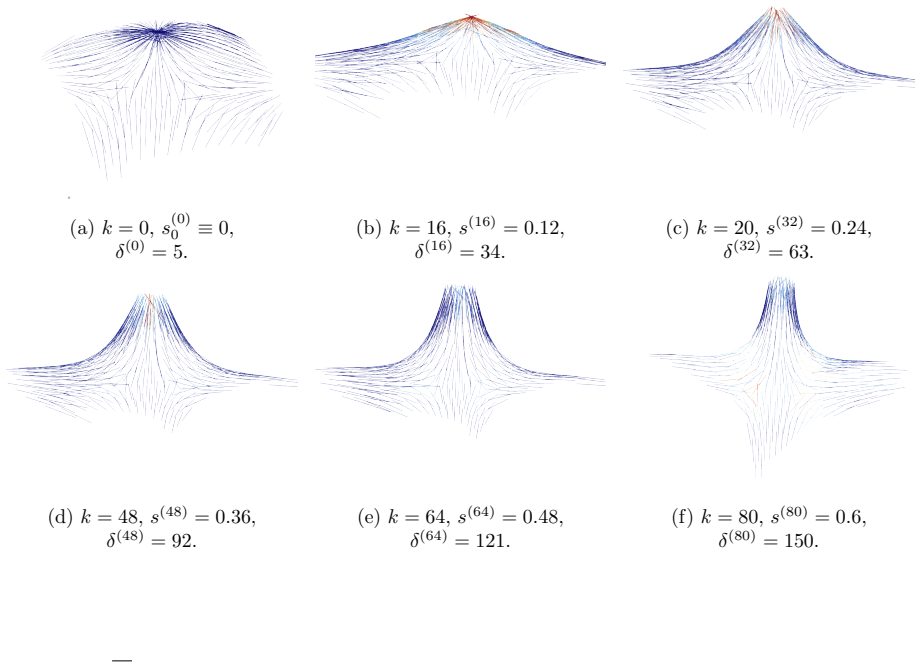
The central claim is that defect topology controls stress localization and shape evolution: +1 defects drive protrusion formation, while -1/2 defects act as structural stabilizers with minimal growth. By varying the initial defect configuration, the model produces diverse morphogenetic outcomes, including uniaxial regeneration, tentacle formation, and biaxial development.
What carries the argument
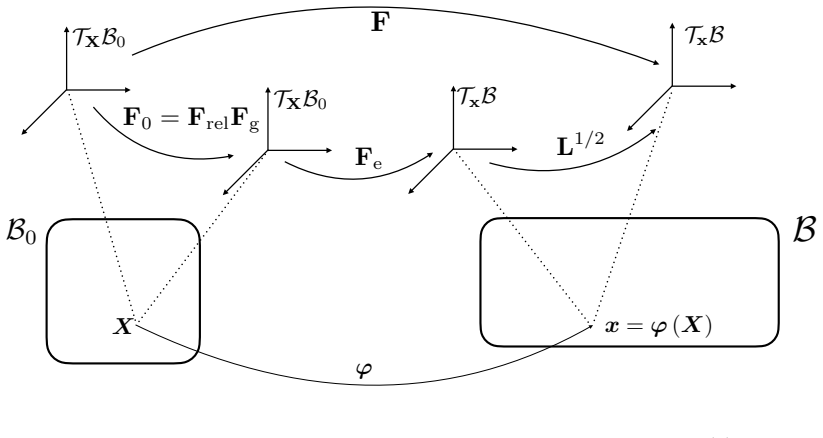
The nematic order parameter together with its topological defects inside a continuum description that couples order to elasticity, stress-driven growth, and adaptive relaxation.
If this is right
- Prescribing +1 defects produces localized protrusions through stress concentration.
- -1/2 defects produce minimal growth and maintain structural stability.
- Different initial defect placements generate uniaxial, tentacled, or biaxial regeneration patterns.
- Heterogeneous growth follows directly from spatial variations in the order parameter.
Where Pith is reading between the lines
- If the mechanism holds, experimental placement of defects could be used to steer regeneration outcomes in living tissues.
- The same stress-defect coupling might operate in other epithelial or cytoskeletal systems that exhibit nematic order.
- A natural extension would test whether defects arise spontaneously or must be seeded to match the observed patterns.
Load-bearing premise
The model assumes that initial defect configurations can be prescribed to match the mature organism's expected morphology and that the resulting quasi-static equilibrium sequence will reproduce observed regeneration outcomes.
What would settle it
Direct imaging showing that +1 defects are absent from future protrusion sites in real Hydra, or that -1/2 defects do not correlate with stable regions, would falsify the claimed control by defect topology.
Figures


read the original abstract
Cellular morphogenesis, the process by which biological tissues acquire shape and structure, remains a fundamental challenge in understanding pattern formation and the coordinated remodeling of cellular assemblies. Under appropriate conditions, cytoskeletal filaments can organize into a nematic phase exhibiting partial orientational order. Topological defects within this nematic organization generate localized mechanical stresses that destabilize the tissue and promote deformation and structural rearrangements to relieve internal stresses. We develop a continuum framework that models living tissues as active biological networks represented as nematic polymer networks capable of heterogeneous growth and remodeling. The model captures macroscopic effects through spatial variations in the fiber order parameter which drives the system away from equilibrium. Morphogenesis is described as a sequence of quasi-static equilibrium states governed by the coupling between nematic order, elasticity, stress-driven growth, and adaptive relaxation. Finite element simulations illustrate Hydra regeneration and development when topological defects are prescribed according to the mature organism's expected morphology. The results show that defect topology controls stress localization and shape evolution: $+1$ defects drive protrusion formation, while $-1/2$ defects act as structural stabilizers with minimal growth. By varying the initial defect configuration, we model diverse morphogenetic outcomes, including uniaxial regeneration, tentacle formation, and biaxial development.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a continuum model treating living tissues as active nematic polymer networks with heterogeneous growth and remodeling. Morphogenesis is represented as a sequence of quasi-static equilibria arising from the coupling of nematic order, elasticity, stress-driven growth, and adaptive relaxation. Finite-element simulations of Hydra regeneration and development are performed with topological defects (+1 and -1/2) prescribed in the initial order-parameter field to match the mature organism's expected morphology; the results indicate that +1 defects localize stress to drive protrusions while -1/2 defects stabilize regions with minimal growth, and that varying the prescribed defect pattern reproduces uniaxial regeneration, tentacle formation, and biaxial outcomes.
Significance. The framework supplies a concrete mechanical link between nematic defect topology and localized growth in a specific biological system. The finite-element implementation permits systematic exploration of different initial defect configurations and their effect on shape evolution. If the defects were instead shown to emerge dynamically from the active-network equations and if the model outputs were compared quantitatively to experimental regeneration data, the approach could strengthen mechanistic understanding of defect-driven morphogenesis.
major comments (3)
- [Abstract] Abstract: the claim that 'defect topology controls stress localization and shape evolution' and that '+1 defects drive protrusion formation' rests on simulations in which initial defect positions are prescribed by hand to match the target mature morphology. This setup demonstrates consistency (prescribed defects produce the expected shapes) but does not establish that defects arise spontaneously from the active-network dynamics without foreknowledge of the final pattern.
- [Abstract] Abstract (model description): the quasi-static equilibrium sequence explicitly decouples defect motion from growth and remodeling. Because defect positions remain fixed inputs rather than evolving quantities, the reported stress localization cannot be attributed to a self-consistent dynamical feedback between activity, order, and shape change.
- [Abstract] Abstract: no validation data, mesh-convergence tests, parameter-sensitivity analysis, or direct quantitative comparison to experimental Hydra regeneration timelines or morphologies are supplied. The central illustrative claim therefore depends on unexamined modeling choices whose robustness is not demonstrated.
minor comments (1)
- [Abstract] Abstract: the statement that the model 'captures macroscopic effects through spatial variations in the fiber order parameter' would benefit from an explicit statement of the free-energy or constitutive relation used for the nematic-elastic coupling.
Simulated Author's Rebuttal
We thank the referee for the constructive and detailed comments. We agree that the model prescribes defect positions and employs a quasi-static approximation without dynamic defect evolution or quantitative experimental validation. We will revise the abstract and add supporting numerical tests to clarify the model's scope and assumptions.
read point-by-point responses
-
Referee: [Abstract] Abstract: the claim that 'defect topology controls stress localization and shape evolution' and that '+1 defects drive protrusion formation' rests on simulations in which initial defect positions are prescribed by hand to match the target mature morphology. This setup demonstrates consistency (prescribed defects produce the expected shapes) but does not establish that defects arise spontaneously from the active-network dynamics without foreknowledge of the final pattern.
Authors: We agree that defects are prescribed in the initial order-parameter field to match expected morphology, as explicitly stated in the abstract. The simulations demonstrate mechanical consistency between prescribed defect topology and resulting shapes but do not address spontaneous emergence. We will revise the abstract to replace 'controls' with 'influences' and to state explicitly that defects are prescribed inputs. revision: yes
-
Referee: [Abstract] Abstract (model description): the quasi-static equilibrium sequence explicitly decouples defect motion from growth and remodeling. Because defect positions remain fixed inputs rather than evolving quantities, the reported stress localization cannot be attributed to a self-consistent dynamical feedback between activity, order, and shape change.
Authors: The referee correctly notes that defect positions are held fixed during the sequence of quasi-static equilibria. This is a deliberate modeling choice to isolate the mechanical consequences of defect topology. We will revise the abstract and model description to emphasize the fixed-defect, quasi-static approximation and its limitations. revision: yes
-
Referee: [Abstract] Abstract: no validation data, mesh-convergence tests, parameter-sensitivity analysis, or direct quantitative comparison to experimental Hydra regeneration timelines or morphologies are supplied. The central illustrative claim therefore depends on unexamined modeling choices whose robustness is not demonstrated.
Authors: We will add mesh-convergence tests and a parameter-sensitivity study for activity strength and relaxation timescales in the revised manuscript. Direct quantitative comparison to experimental regeneration timelines is not performed in the current work, which remains qualitative; we will note this as a limitation and direction for future study. revision: partial
Circularity Check
Defect positions prescribed to match target morphology make outcomes by construction
specific steps
-
fitted input called prediction
[Abstract]
"Finite element simulations illustrate Hydra regeneration and development when topological defects are prescribed according to the mature organism's expected morphology. The results show that defect topology controls stress localization and shape evolution: +1 defects drive protrusion formation, while -1/2 defects act as structural stabilizers with minimal growth. By varying the initial defect configuration, we model diverse morphogenetic outcomes, including uniaxial regeneration, tentacle formation, and biaxial development."
Defect configurations are chosen to match the target morphology in advance; the reported shape evolution is then the direct output of evolving equilibria under that fixed prescription, so the claimed control by defect topology reduces to a consistency check on the inputs rather than an independent derivation.
full rationale
The central claim that defect topology controls morphogenesis is demonstrated only after the model explicitly sets initial defect locations by hand to reproduce the mature organism's expected morphology. The subsequent quasi-static evolution then necessarily produces the corresponding shapes (protrusions at +1 defects, stabilization at -1/2), rendering the reported control a direct consequence of the prescribed inputs rather than an emergent prediction from the active network dynamics. This matches the fitted-input-called-prediction pattern and accounts for the observed partial circularity; the remainder of the continuum framework appears self-contained.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
Bladon P, Terentjev EM, Warner M. 1994. Deformation-induced orientational tran- sitions in liquid crystals elastomer, Journal of Physics II France 4(1), 75–91 (doi:10.1051/jp2:1994100)
-
[2]
Bailles A, Serafini G, Andreas H, Zechner C, Modes CD, Tomancak P. 2025. Anisotropic stretch biases the self-organization of actin fibers in multicellular Hydra ag- gregates, Proceedings of the National Academy of Sciences 122(32), e2423437122 (doi: 10.1073/pnas.2423437122)
-
[3]
Blanch-Mercader C, Yashunsky V, Garcia S, Duclos G, Giomi L, Silberzan P. 2018. Tur- bulent dynamics of epithelial cell cultures, Physical Review Letters 120, 208101 (doi: 10.1103/PhysRevLett.120.208101)
-
[4]
Doostmohammadi A, Ign´ es-Mullol J, Yeomans JM, Sagu’es F. 2018. Active nematics, Na- ture Communication 9, 3246 (doi: 10.1038/s41467-018-05666-8)
-
[5]
Ericksen JL. 1991. Liquid crystals with variable degree of orientations, Archive for Rational Mechanics and Analysis 113, 97-120 (doi:10.1007/BF00380413)
-
[6]
Goodbrake C, Goriely A, Yavari A. 2021. The mathematical foundations of anelasticity: existence of smooth global intermediate configurations, Proceedings of the Royal Society A 477, 20200462 (doi: 10.1098/rspa.2020.0462)
-
[7]
Goriely A. 2017. The Mathematics and Mechanics of Biological Growth, Springer-Verlag, New York. 36
2017
-
[8]
Guillamat P, Mirza W, Bal PK, G´ omez-Gomz´ ales M, Roca-Cusachs P, Arroyo M, Trepat X. 2026. Guidance of cellular nematic elastomers into shape-programmable living surfaces, Science 392, 317-323 (doi: 10.1126/science.adz9174)
-
[9]
Hernandez A, Zhu C, Giomi L. 2026. Mechanics of axis formation in Hydra, arXiv a2601.05220 (doi:arxiv.org/abs/2601.05220)
arXiv 2026
-
[10]
Hoffmann LA, Carenza LN, Eckert J, Giomi L. 2022. Theory of defect-mediated morpho- genesis, Science Advances 8, eabk2712 (doi:10.1126/sciadv.abk2712)
-
[11]
Hoffmann LA, Carenza LN, Giomi L. 2023. Tuneable defect-curvature coupling and topo- logical transitions in active shells, Soft Matter 19, 3423–3435 (doi:10.1039/D3SM00071K)
-
[12]
Hopf H. 1983. Differential Geometry in the Large, Springer-Verlag, Berlin, Heidelberg
1983
-
[13]
Huang R, Ogden RW, Penta R. 2025. Modelling volumetric growth in soft solids via residual stress, Journal of Elasticity 157(4), 73 (doi: 10.1007/s10659-025-10164-2)
-
[14]
Kawaguchi K, Kageyama R, Sano M. 2017. Topological defects control collective dynamics in neural progenitor cell cultures, Nature 545, 212 (doi: 10.1038/nature22321)
-
[15]
Copenhagen K, Alert R, Wingreen N, Shaevitz J. 2021. Topological defects promote layer formation in myxococcusxanthus colonies, Nature Physics 17, 211 (doi: 10.1038/s41567- 020-01056-4)
-
[16]
Keber FC, Loiseau E, Sanchez T, DeCamp SJ, Giomi L, Bowick MJ, Marchetti MC, Dogic Z, Bausch AR. 2014. Topology and dynamics of active nematic vesicles, Science 345 (6201), 1135-1139 (doi: 10.1126/science.1254784)
-
[17]
Kumar N, Zhang R, de Pablo JJ, Gardel ML. 2018. Tunable structure and dynamics of active liquid crystals, Science Advances 4, eaat7779 (doi: 10.1126/sciadv.aat7779)
-
[18]
Lee P, Wolgemuth CW. 2011. Crawling cells can close wounds without purse strings or sig- naling, PLOS Computational Biology 7(3), e1002007 (doi: 10.1371/journal.pcbi.1002007)
-
[19]
Livshits A, Shani-Zerbib L, Maroudas-Sacks Y. 2017. Structural inheritance of the actin cytoskeletal organization determines the body axis in regenerating Hydra, Cell Reports 18, 1410-1421 (doi: 10.1016/j.celrep.2017.01.036)
-
[20]
Lubarda VA. 2004. Constitutive theories based on the multiplicative decomposition of de- formation gradient: thermoelasticity, elastoplasticity and biomechanics, Applied Mechanics Reviews 57(2), 95-108 (doi: 10.1115/1.1591000)
-
[21]
Marchetti MC, Joanny JF, Ramaswamy S, Liverpool TB, Prost J, Rao M, Simha RA
-
[22]
Hydrodynamics of soft active matter, Reviews of Modern Physics 85, 1143 (doi: 10.1103/RevModPhys.85.1143)
-
[23]
Maroudas-Sacks Y, Garion L, Shani-Zerbib L, Livshits A, Braun E, Keren K. 2020. Topo- logical defects in the nematic order of actin fibers as organization centers of Hydra mor- phogenesis, Nature Physics 17(2), 1-9 (doi: 10.1101/2020.03.02.972539)
-
[24]
Maroudas-Sacks Y, Suganthan S, Garion L, Ascoli-Abbina Y, Westfried A, Dori N, Pasv- inter I, Popovi´ c M, Keren K. 2025. Mechanical strain focusing at topological defect sites in regenerating Hydra, Development 152(4) (doi: 10.1242/dev.204514). 37
-
[25]
Mihai LA. 2022. Stochastic Elasticity: A Nondeterministic Approach to the Nonlinear Field Theory, Springer Cham, Switzerland (doi: 10.1007/978-3-031-06692-4)
-
[26]
Napoli G, Paparini S. 2016. Nematic bubbles and the breaking of spherical symmetry, arXiv 2603.07082 (doi: arxiv.org/abs/2603.07082)
arXiv 2016
-
[27]
Narayan V, Ramaswamy S, Menon N. 2007. Long-lived giant number fluctuations in a swarming granular nematic, Science 317, 105 (doi: 10.1126/science.1140414)
-
[28]
Paparini S, Giusteri GG, Mihai LA. 2025. Shape instabilities driven by topological defects in nematic polymer networks, Journal of Elasticity 157, 69 (doi: 10.1007/s10659-025-10160-6)
-
[29]
Pearce DJG, Thibault C, Chaboche Q, Blanch-Mercader C. 2025. Passive defect driven morphogenesis in nematic membranes, Physical Review Letters 134(1), 018402 (doi: 10.1103/PhysRevLett.134.018402)
-
[30]
P´ erez-Gonz´ alez C, Alert R, Blanch-Mercader C, G´ omez-Gonz´ alez M, Kolodziej T, Bazel- lieres E, Casademunt J, Trepat X. 2019. Active wetting of epithelial tissues, Nature Physics 15(1), 79-88 (doi: 10.1038/s41567-018-0279-5)
-
[31]
Perros T, Biquet-Bisquert A, Ben Meriem Z, Delarue M, Joseph P, Marcq P, Cochet- Escartin O. 2025. Mechanical characterization of regenerating Hydra tissue spheres, Bio- physical Journal 123, 1792-1803 (doi: 10.1016/j.bpj.2024.05.022)
-
[32]
Poincar´ e, H. 1886. Sur les courbes d´ efinies par les ´ equations diff´ erentielles (quatri` eme par- tie), Journal de Math´ ematiques Pures et Appliqu´ ees, 4e s´ erie, 2, 151–217
-
[33]
Ravichandran Y, Vogg M, Kruse K, Pearce DJG, Roux A. 2025. Topology changes of Hydra define actin orientation defects as organizers of morphogenesis, Science Advances 11(3), eadr9855 (doi: 10.1126/sciadv.adr98)
-
[34]
Rosso R, Virga EG, Kralj S. 2012. Parallel transport and defects on nematic shells, Con- tinuum Mechanics and Thermodynamics 24, 643-664 (doi:10.1007/s00161-012-0259-4)
-
[35]
Sadik S, Yavari A. 2017. On the origins of the idea of the multiplicative decomposition of the deformation gradient, Mathematics and Mechanics of Solids 22(4), 771-772 (doi: 10.1177/1081286515612280)
-
[36]
Saw TB, Doostmohammadi A, Nier V, Kocgozlu L, Thampi S, Toyama Y, Marcq P, Lim C, Yeomans J, Ladoux B. 2017. Topological defects in epithelia govern cell death and extrusion, Nature 544, 212 (doi: 10.1038/nature21718)
-
[37]
Stoker JJ. 1989. Differential Geometry, Wiley, Wiley Classics Library
1989
-
[38]
Wang Z, Marchetti MC, Brauns F. 2023. Patterning of morphogenetic anisotropy fields. Proceedings of the National Academy of Sciences 120, e2220167120 (doi: 10.1073/pnas.2220167120)
-
[39]
Warner M, Gelling KP, Vilgis TA. 1988. Theory of nematic networks, The Journal of Chemical Physics 88(6), 4008–4013 (doi:10.1063/1.453852)
-
[40]
Warner M, Terentjev EM. 2007. Liquid Crystal Elastomers, paper back, Oxford University Press, Oxford, UK
2007
-
[41]
Warner M, Wang X. 1991. Elasticity and phase behavior of nematic elastomers, Macro- molecules 24, 4932-4941 (doi: 10.1021/ma0 0 017a033). 38
work page doi:10.1021/ma0 1991
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.