Sharpness characterizes Hill functions
Pith reviewed 2026-06-27 12:07 UTC · model grok-4.3
The pith
Rational functions with 0 ≤ α_i ≤ β_i have sharpness at most n/4, achieved exactly by Hill functions of degree n.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Any rational function r(x)=(α0+α1x+⋯+αn xn)/(β0+β1x+⋯+βn xn) with real coefficients 0≤αi≤βi has sharpness at most n/4, with equality if and only if r is a Hill function with Hill coefficient n.
What carries the argument
The pointwise coefficient constraint 0 ≤ α_i ≤ β_i on the rational function, which forces the semi-log derivative to stay below the value n/4.
If this is right
- Hill functions of degree n are the unique maximizers of sharpness under the given coefficient constraints.
- No other rational function meeting the inequalities can exceed sharpness n/4.
- The result supplies the precise mathematical content of the Hopfield barrier for sharpness in energy-free biological computation.
Where Pith is reading between the lines
- If real biological maps are close to these rational forms, natural selection would be expected to converge on Hill-like responses for any task requiring the steepest possible switch.
- The same bounding technique may apply to other response metrics such as integrated sensitivity or noise robustness.
- Synthetic circuit design could use the equality case to guarantee maximal sharpness without increasing molecular complexity beyond degree n.
Load-bearing premise
Biological input-output maps are adequately modeled by rational functions whose numerator and denominator coefficients satisfy 0 ≤ α_i ≤ β_i for every i.
What would settle it
Exhibit one rational function of degree n obeying 0 ≤ α_i ≤ β_i whose semi-log derivative somewhere exceeds n/4.
Figures

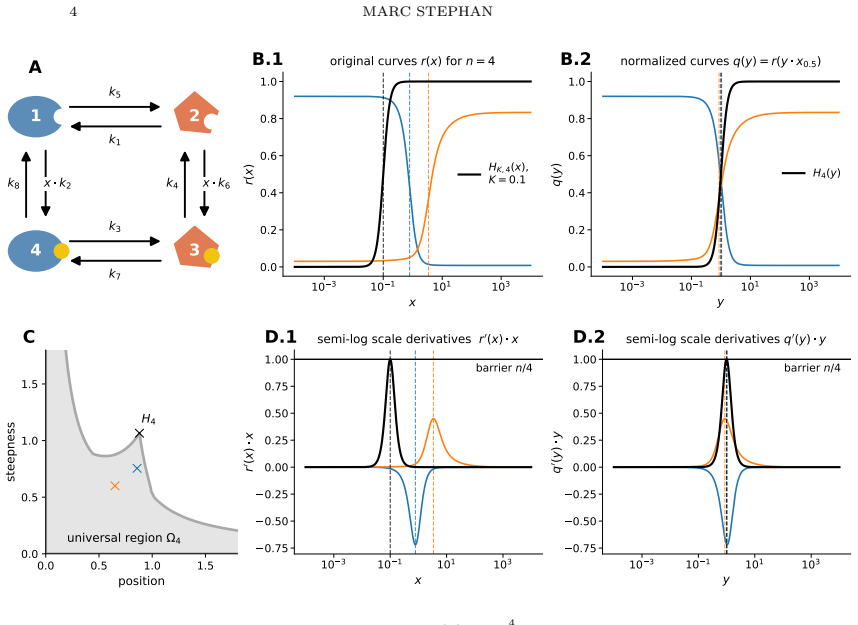
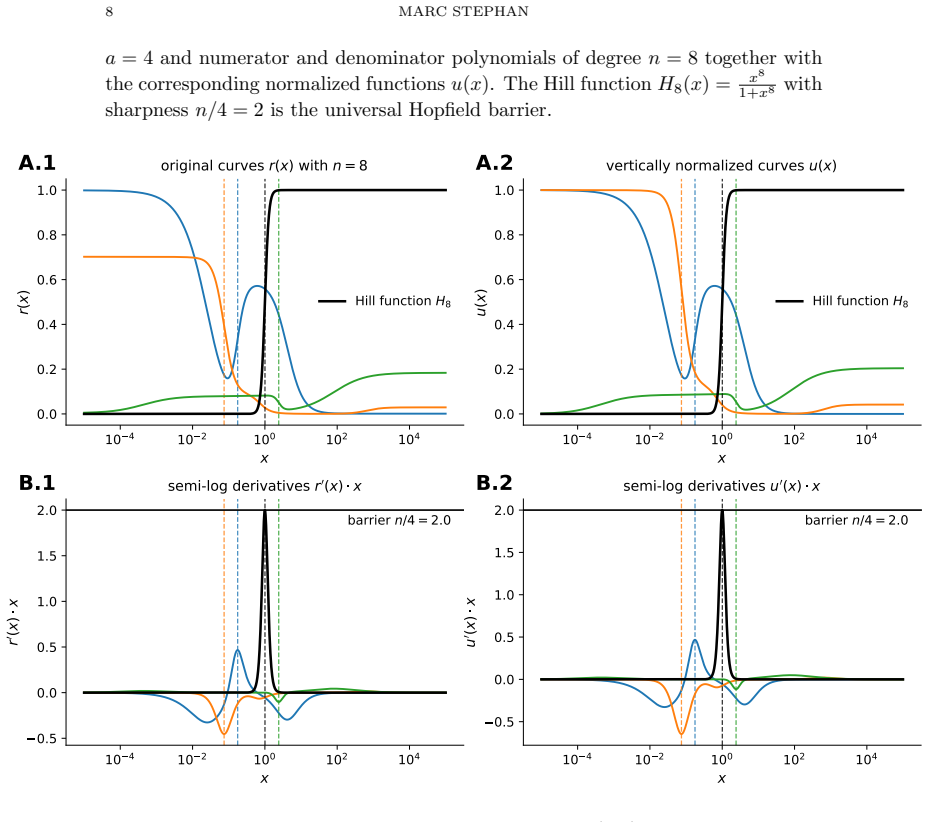
read the original abstract
While long treated as empirical fits, Hill functions have been postulated to be the universal Hopfield barrier for sharpness of input-output responses by Martinez-Corral, Nam, DePace, and Gunawardena. A Hopfield barrier is a fundamental limit on how well biological systems can process information without expending energy. Their case rested on numerical findings for Hill coefficients $4$ and $6$. We give a precise formulation and proof of this: measuring sharpness by the supremum of the derivative in semi-log scale, any rational function $r(x)=(\alpha_0+\alpha_1 x+ \cdots +\alpha_n x^n)/(\beta_0 + \beta_1 x+ \cdots + \beta_n x^n)$ with real coefficients $0\leq \alpha_i\leq \beta_i$ has sharpness at most $n/4$, with equality if and only if $r$ is a Hill function with Hill coefficient $n$.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript provides a precise formulation and proof that any rational function r(x) = (α₀ + α₁x + ⋯ + αₙxⁿ)/(β₀ + β₁x + ⋯ + βₙxⁿ) with real coefficients satisfying 0 ≤ α_i ≤ β_i for all i has sharpness (sup_{x>0} x r'(x)) at most n/4, with equality if and only if r is the Hill function of coefficient n. This is framed as establishing the Hopfield barrier for sharpness of input-output maps in the biological setting.
Significance. If the central theorem holds, the result supplies a parameter-free mathematical proof of a fundamental limit on sharpness for a broad class of rational input-output maps, confirming and generalizing the numerical observations of Martinez-Corral et al. for Hill coefficients 4 and 6. The if-and-only-if characterization of equality cases is a strong feature, as is the derivation directly from the coefficient inequalities without additional assumptions or fitted parameters.
minor comments (2)
- The abstract and introduction should explicitly state the precise definition of the semi-log derivative (i.e., whether sharpness is sup x r'(x) or sup (x r'(x)/r(x)) or another variant) to avoid any ambiguity for readers outside the immediate subfield.
- Section stating the Hopfield barrier formulation would benefit from a short remark clarifying why the coefficient condition 0 ≤ α_i ≤ β_i is the natural modeling assumption for the biological input-output maps under consideration.
Simulated Author's Rebuttal
We thank the referee for their positive assessment of the manuscript and their recommendation to accept. No revisions are required.
Circularity Check
No significant circularity; derivation is algebraic proof from stated assumptions
full rationale
The central result is a mathematical bound on sharpness (sup x r'(x)) for rational functions of degree n satisfying 0 ≤ α_i ≤ β_i. This follows directly from the functional form and coefficient inequalities without any fitted parameters, self-referential definitions, or load-bearing self-citations. The equality case for Hill functions is shown by direct verification within the same algebraic framework. No step reduces the claimed prediction to an input by construction.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption Biological input-output maps belong to the class of rational functions with 0 ≤ α_i ≤ β_i
- domain assumption Sharpness is the supremum of the derivative of log r with respect to log x
Reference graph
Works this paper leans on
-
[1]
, title =
Popoviciu, T. , title =. Mathematica, Cluj , volume =. 1935 , language =
1935
-
[2]
Bhatia, Rajendra and Davis, Chandler , TITLE =. Amer. Math. Monthly , FJOURNAL =. 2000 , NUMBER =. doi:10.2307/2589180 , URL =
-
[3]
Muilwijk , journal =
J. Muilwijk , journal =. Note on a
-
[4]
Hill, A. V. The possible effects of the aggregation of the molecules of haemoglobin on its dissociation curves. J Physiol (Lond). 1910
1910
-
[5]
The linear framework: using graph theory to reveal the algebra and thermodynamics of biomolecular systems , volume =. Interface Focus , author =. 2022 , pages =. doi:10.1098/rsfs.2022.0013 , abstract =
-
[6]
and Gunawardena, Jeremy , month = may, year =
Martinez-Corral, Rosa and Nam, Kee-Myoung and DePace, Angela H. and Gunawardena, Jeremy , month = may, year =. The. Proceedings of the National Academy of Sciences , publisher =. doi:10.1073/pnas.2318329121 , abstract =
-
[7]
The. The FASEB Journal , author =. 1997 , note =. doi:10.1096/fasebj.11.11.9285481 , abstract =
-
[8]
Nam, Kee-Myoung and Gunawardena, Jeremy , month = nov, year =. The linear framework. Frontiers in Cell and Developmental Biology , publisher =. doi:10.3389/fcell.2023.1233808 , abstract =
-
[9]
Motulsky, Harvey and Christopoulos, Arthur , month = may, year =. Fitting. doi:10.1093/oso/9780195171792.001.0001 , abstract =
-
[10]
A hundred years of the. The Biochemist , author =. 2013 , pages =. doi:10.1042/BIO03506040 , abstract =
-
[11]
Santillán, M. , year =. On the. Mathematical Modelling of Natural Phenomena , publisher =. doi:10.1051/mmnp:2008056 , abstract =
-
[12]
Archive for History of Exact Sciences , author =
The. Archive for History of Exact Sciences , author =. 2012 , keywords =. doi:10.1007/s00407-012-0098-5 , abstract =
-
[13]
Estrada, Javier and Wong, Felix and DePace, Angela and Gunawardena, Jeremy , month = jun, year =. Information. Cell , publisher =. doi:10.1016/j.cell.2016.06.012 , abstract =
-
[14]
Proceedings of the National Academy of Sciences of the United States of America , author =
Kinetic proofreading: a new mechanism for reducing errors in biosynthetic processes requiring high specificity , volume =. Proceedings of the National Academy of Sciences of the United States of America , author =. 1974 , keywords =. doi:10.1073/pnas.71.10.4135 , abstract =
-
[15]
Owen, Jeremy A. and Horowitz, Jordan M. , month = mar, year =. Size limits the sensitivity of kinetic schemes , volume =. Nature Communications , publisher =. doi:10.1038/s41467-023-36705-8 , abstract =
-
[16]
Martinez-Corral, Rosa , month = may, year =. rosamc/universal-boundaries-. doi:10.5281/zenodo.11119219 , publisher =
-
[17]
Allosteric conformational ensembles have unlimited capacity for integrating information , volume =
Biddle, John W and Martinez-Corral, Rosa and Wong, Felix and Gunawardena, Jeremy , editor =. Allosteric conformational ensembles have unlimited capacity for integrating information , volume =. eLife , publisher =. 2021 , keywords =. doi:10.7554/eLife.65498 , abstract =
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.