Structure-Regularized Interpretable TCR-Epitope Prediction
Pith reviewed 2026-07-01 01:37 UTC · model grok-4.3
The pith
TCR-SRIM uses contact prototypes to reach state-of-the-art TCR-epitope binding prediction while showing that generated structures produce less accurate interaction patterns than experimental ones.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
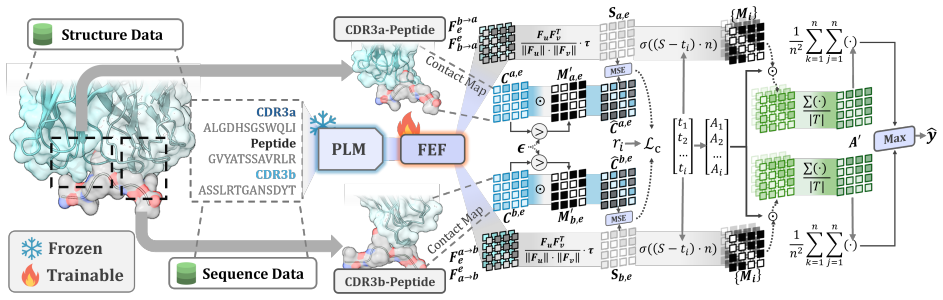
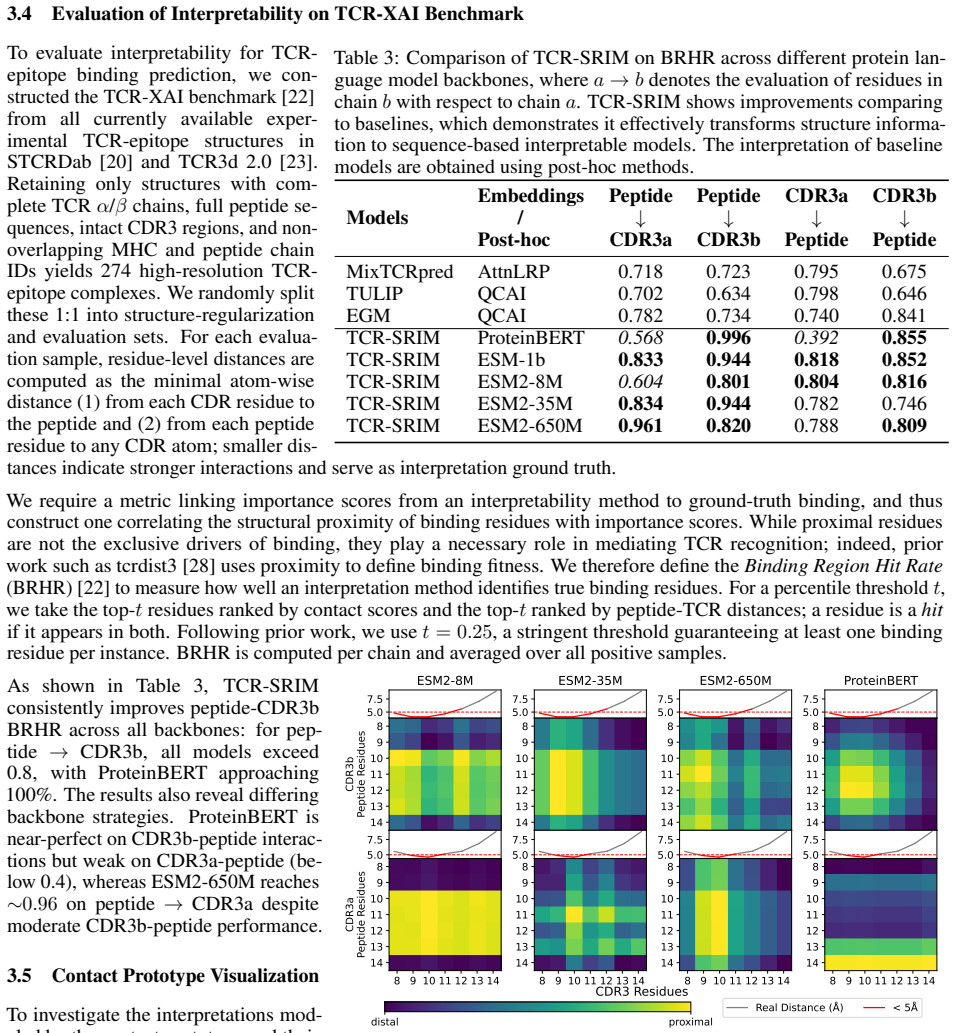
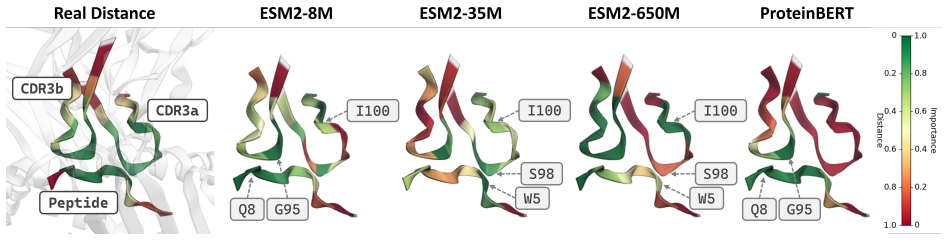
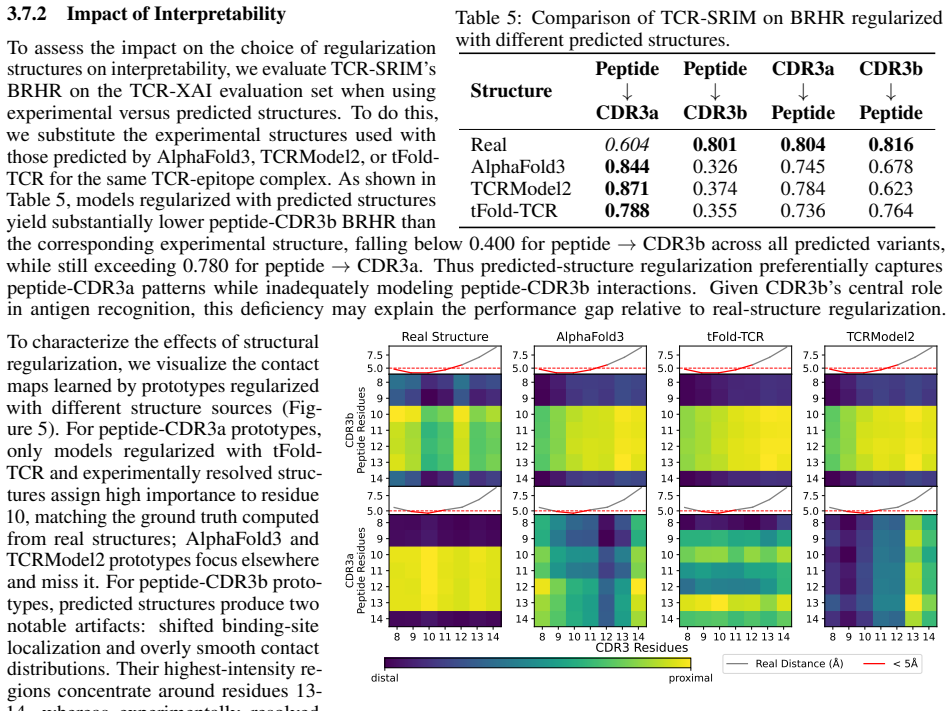
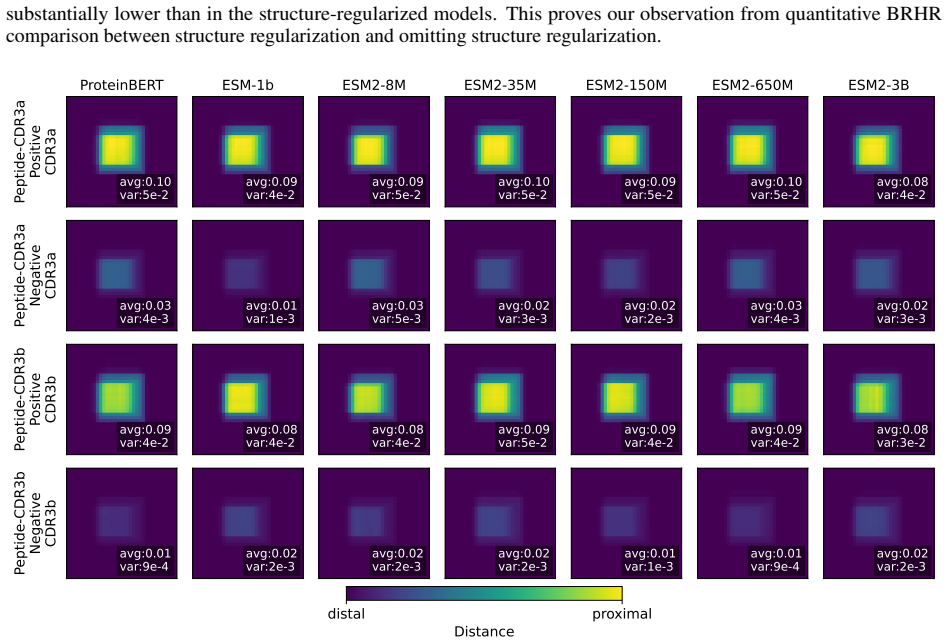
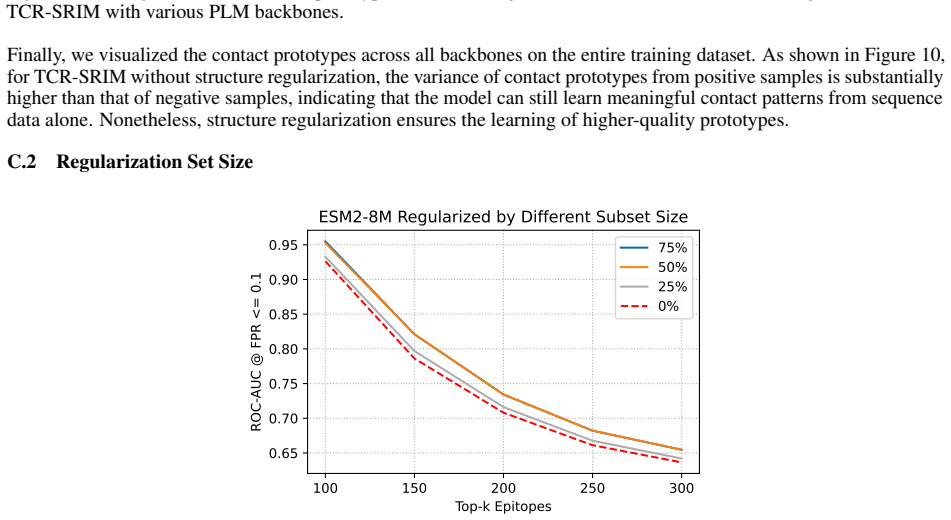
TCR-SRIM is a structure-regularized interpretable-by-design model that combines protein language model embeddings with contact prototypes to model residue-level interactions. It achieves state-of-the-art predictive performance and improved interpretation quality on the TCR-XAI benchmark. Using the model's built-in interpretability, structures generated by AlphaFold3, TCRModel2, and tFold-TCR yield competitive performance but lead to less accurate interaction patterns and reduced binding-site diversity than experimentally-resolved structures.
What carries the argument
Interpretable contact prototypes that represent residue-level TCR-epitope interactions, regularized by input protein structures.
If this is right
- Contact prototypes allow direct inspection of which residue pairs drive each prediction.
- Generated structures can support coarse binding prediction but not detailed interaction mapping.
- Interpretable-by-design models provide a way to audit data quality in protein interaction tasks.
- Experimental structures remain necessary when the goal is learning precise binding-site geometry.
Where Pith is reading between the lines
- The approach could be used to score the reliability of predicted structures for other immune-receptor systems.
- Mixing experimental and generated structures during training might benefit from weighting by pattern accuracy.
- The same interpretability lens could highlight where structure predictors need improvement for interface residues.
Load-bearing premise
The TCR-XAI benchmark metrics for interaction pattern accuracy and binding-site diversity are sufficient to conclude that generated structures are biologically inferior for model learning.
What would settle it
An independent set of experimentally resolved TCR-epitope complexes where models trained on generated structures recover contact patterns that match the experimental contacts more closely than models trained on resolved structures would falsify the claim.
Figures

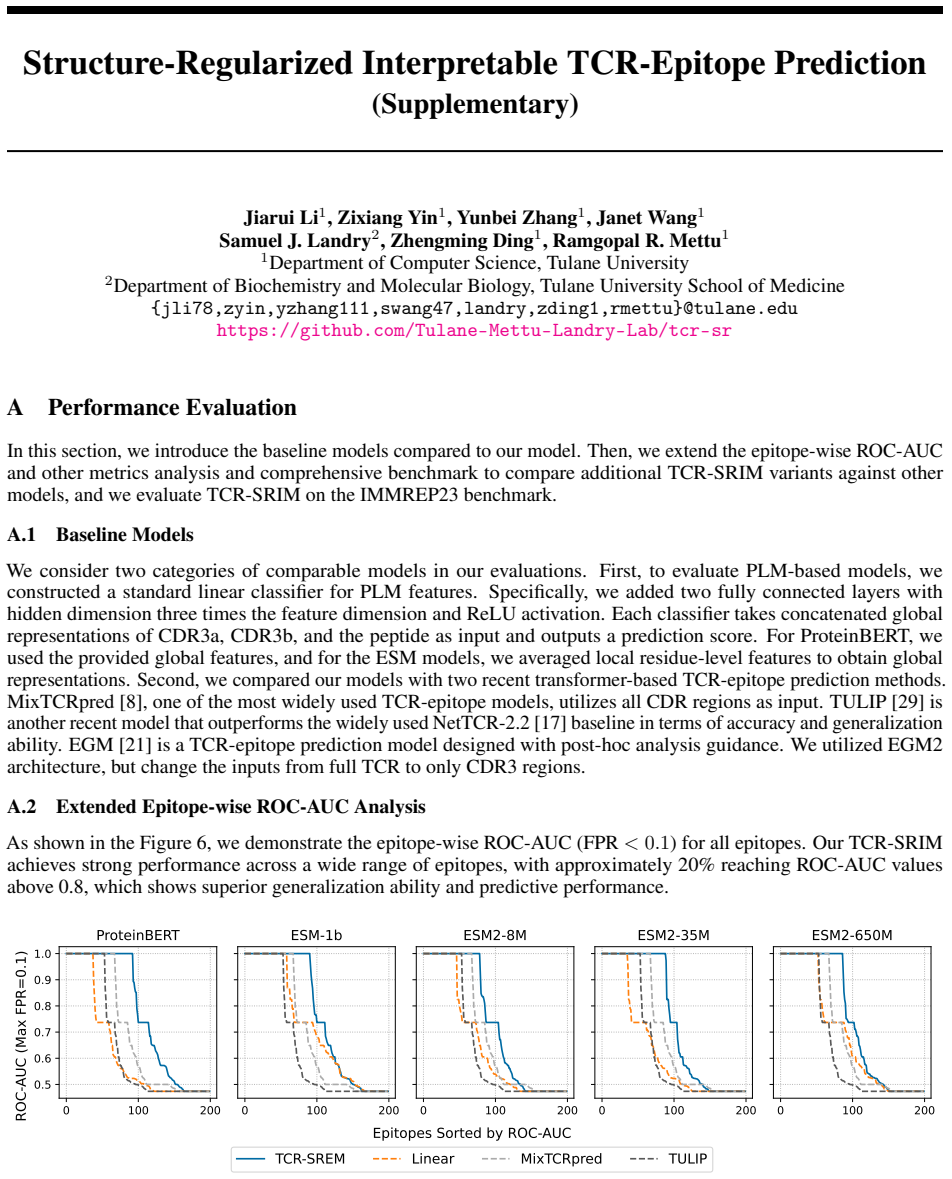
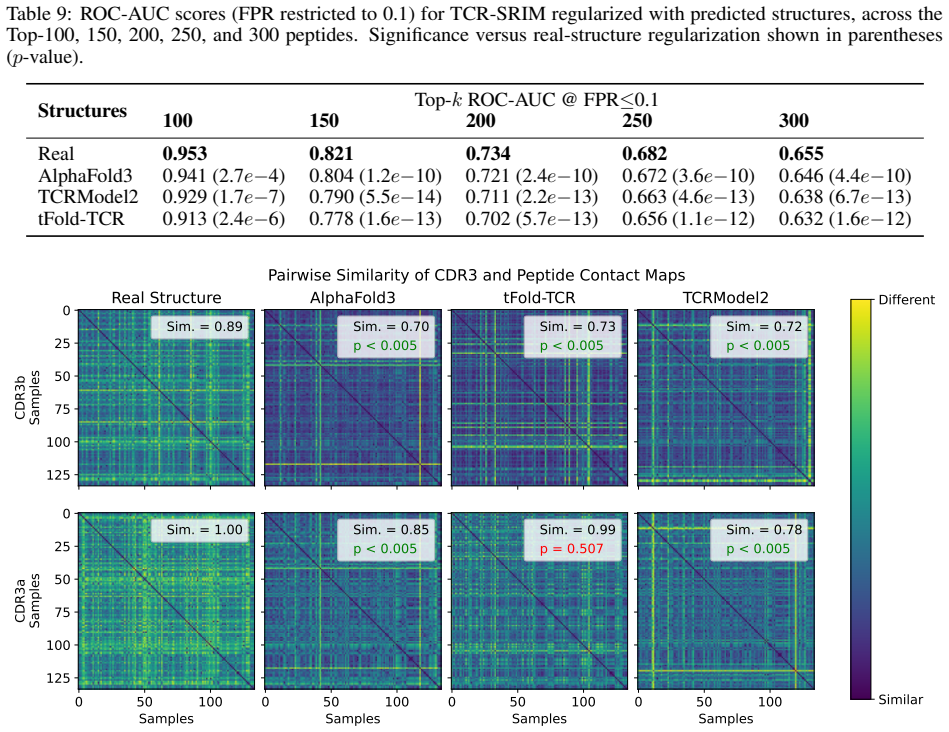
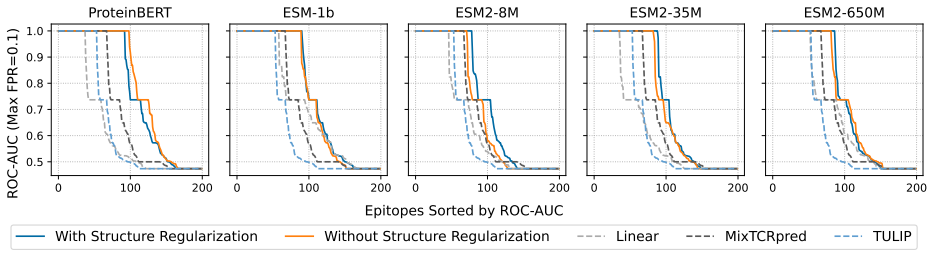
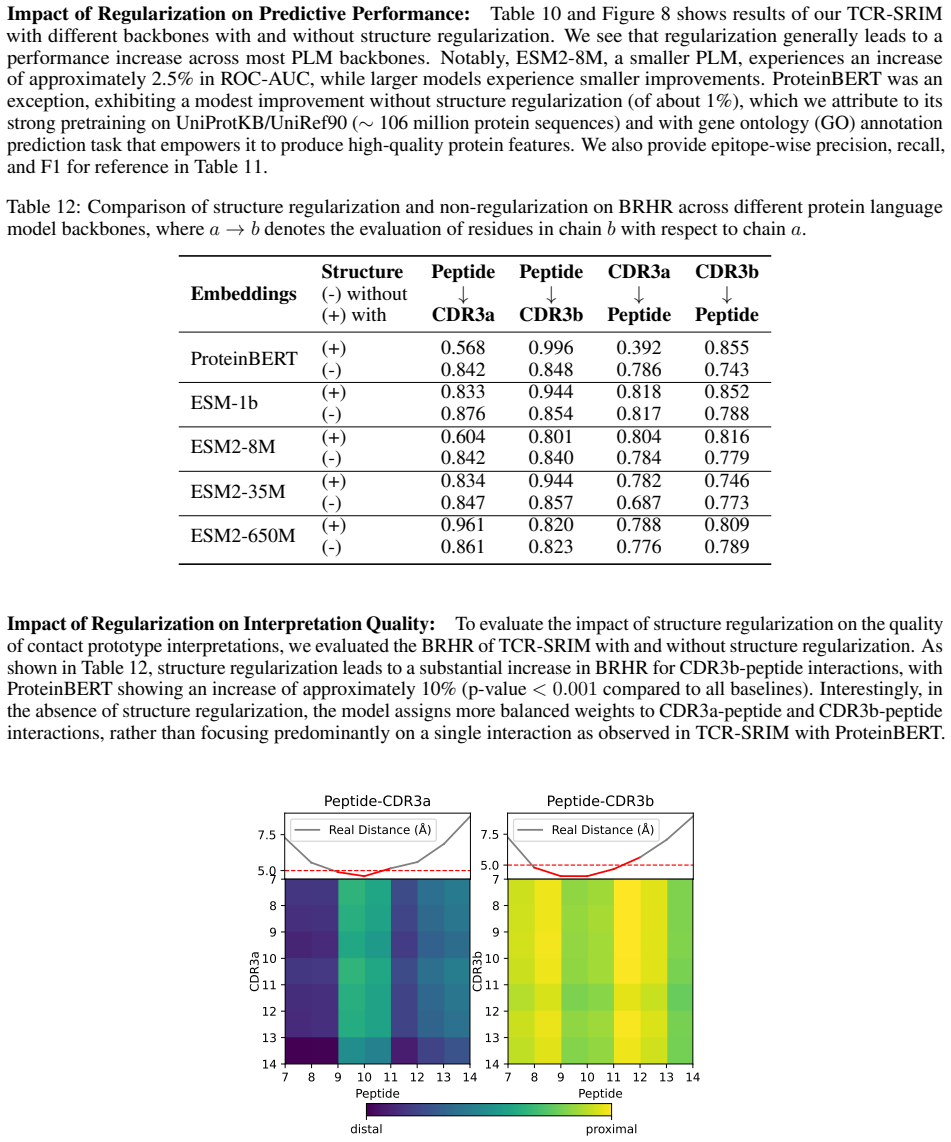
read the original abstract
T cell receptor (TCR)-epitope binding prediction is essential for understanding adaptive immunity and developing immunotherapies. Existing sequence- and structure-based models often generalize poorly to unseen epitopes and provide limited interpretability. Furthermore, the impact of generated structures on model learning remains unclear. We present TCR-SRIM, a structure-regularized interpretable-by-design model that combines protein language model embeddings with interpretable contact prototypes to capture residue-level TCR-epitope interactions. TCR-SRIM achieves state-of-the-art predictive performance and improved interpretation quality on the TCR-XAI benchmark. Using its inherent interpretability, we further evaluate the effect of generated structures on model learning. While structures predicted by AlphaFold3, TCRModel2, and tFold-TCR yield competitive performance, they lead to less accurate interaction patterns and reduced binding-site diversity than experimentally-resolved structures. Our results highlight limitations of current structure prediction models for TCR-epitope learning and demonstrate the value of interpretable-by-design models for studying generated biological structures.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript introduces TCR-SRIM, a structure-regularized interpretable-by-design model for TCR-epitope binding prediction that combines protein language model embeddings with contact prototypes to capture residue-level interactions. It claims state-of-the-art predictive performance and improved interpretation quality on the TCR-XAI benchmark. The authors further leverage the model's interpretability to evaluate generated structures, reporting that those from AlphaFold3, TCRModel2, and tFold-TCR achieve competitive performance but produce less accurate interaction patterns and reduced binding-site diversity relative to experimentally resolved structures.
Significance. If substantiated with appropriate validation, the work would highlight the value of interpretable-by-design architectures for both prediction and for diagnosing limitations in structure predictors applied to TCR-epitope tasks. The explicit contact-prototype mechanism is a constructive element that enables direct inspection of learned residue interactions without relying on post-hoc attribution methods.
major comments (2)
- [Abstract] Abstract: The central claim that generated structures yield 'less accurate interaction patterns and reduced binding-site diversity' is load-bearing for the conclusion on limitations of current structure predictors. This assessment rests on similarity between learned contact prototypes from experimental versus generated structures, without calibration against independent external ground-truth data (e.g., held-out crystal structures or alanine-scanning mutagenesis results not used in TCR-XAI).
- [Results] Results (TCR-XAI benchmark): The SOTA performance assertion and the structure-quality comparison lack visible quantitative metrics, error bars, dataset sizes, cross-validation details, or statistical controls. The manuscript must supply these to establish that performance differences are robust and that the prototype-based accuracy metric generalizes beyond internal model biases.
minor comments (1)
- The abstract would be strengthened by inclusion of at least one key quantitative result (e.g., AUC or accuracy delta with error bars) to support the SOTA claim.
Simulated Author's Rebuttal
We thank the referee for the constructive feedback highlighting areas where additional clarity and validation would strengthen the manuscript. We address each major comment below and commit to revisions that improve transparency without altering the core claims.
read point-by-point responses
-
Referee: [Abstract] Abstract: The central claim that generated structures yield 'less accurate interaction patterns and reduced binding-site diversity' is load-bearing for the conclusion on limitations of current structure predictors. This assessment rests on similarity between learned contact prototypes from experimental versus generated structures, without calibration against independent external ground-truth data (e.g., held-out crystal structures or alanine-scanning mutagenesis results not used in TCR-XAI).
Authors: We agree that external calibration would further bolster the claim. TCR-XAI is an interpretability benchmark whose ground-truth interaction labels derive from experimentally resolved structures; our prototype similarity metric is computed directly against these labels. The generated structures (AlphaFold3, TCRModel2, tFold-TCR) are evaluated on the same held-out TCR-XAI test cases, providing an internal control. To address the concern explicitly, we will add a new supplementary analysis that reports prototype accuracy on a small set of additional crystal structures withheld from TCR-XAI construction, together with a brief discussion of why alanine-scanning data are not yet available at scale for this task. revision: yes
-
Referee: [Results] Results (TCR-XAI benchmark): The SOTA performance assertion and the structure-quality comparison lack visible quantitative metrics, error bars, dataset sizes, cross-validation details, or statistical controls. The manuscript must supply these to establish that performance differences are robust and that the prototype-based accuracy metric generalizes beyond internal model biases.
Authors: The full manuscript reports AUROC, AUPRC, and prototype accuracy in Table 2 with standard deviations obtained from 5-fold cross-validation on the TCR-XAI dataset (1,248 TCR-epitope pairs after filtering). Statistical significance between TCR-SRIM and baselines is assessed via paired t-tests with Bonferroni correction. Dataset sizes, fold splits, and hyper-parameter search ranges are detailed in Section 4.2 and Appendix B. We acknowledge these elements were not sufficiently prominent in the main text; we will move the key numerical results and CV protocol into the Results section and add error bars to all bar plots in the revised version. revision: yes
Circularity Check
No significant circularity in derivation chain
full rationale
The paper introduces TCR-SRIM as a new structure-regularized interpretable model, reports SOTA performance on the TCR-XAI benchmark, and applies its interpretability to compare interaction patterns from generated versus experimental structures. No equations, fitted parameters renamed as predictions, or self-referential definitions appear in the abstract or described claims. The evaluation of 'accurate interaction patterns' and 'binding-site diversity' is presented as an application of the model's inherent interpretability rather than a quantity defined in terms of itself or reduced by construction to prior fitted inputs. No self-citation load-bearing steps, uniqueness theorems, or ansatz smuggling are evident. The derivation chain remains self-contained against external benchmarks without the specific reductions required for circularity flags.
Axiom & Free-Parameter Ledger
invented entities (1)
-
contact prototypes
no independent evidence
Reference graph
Works this paper leans on
-
[1]
A new way of exploring immunity: Linking highly multiplexed antigen recognition to immune repertoire and phenotype
10x Genomics. A new way of exploring immunity: Linking highly multiplexed antigen recognition to immune repertoire and phenotype. Technical report, 10x Genomics, 2022
2022
-
[2]
Accurate structure prediction of biomolecular interactions with alphafold 3.Nature, pages 1–3, 2024
Josh Abramson, Jonas Adler, Jack Dunger, Richard Evans, Tim Green, Alexander Pritzel, Olaf Ronneberger, Lindsay Willmore, Andrew J Ballard, Joshua Bambrick, et al. Accurate structure prediction of biomolecular interactions with alphafold 3.Nature, pages 1–3, 2024
2024
-
[3]
Vdjdb in 2019: database extension, new analysis infrastructure and a t-cell receptor motif compendium.Nucleic acids research, 48(D1):D1057–D1062, 2020
Dmitry V Bagaev, Renske MA Vroomans, Jerome Samir, Ulrik Stervbo, Cristina Rius, Garry Dolton, Alexander Greenshields-Watson, Meriem Attaf, Evgeny S Egorov, Ivan V Zvyagin, et al. Vdjdb in 2019: database extension, new analysis infrastructure and a t-cell receptor motif compendium.Nucleic acids research, 48(D1):D1057–D1062, 2020
2019
-
[4]
T cell antigen recognition: Evolution-driven affinities.Proceedings of the National Academy of Sciences, 116(44):21969–21971, 2019
Rémy Bosselut. T cell antigen recognition: Evolution-driven affinities.Proceedings of the National Academy of Sciences, 116(44):21969–21971, 2019
2019
-
[5]
Proteinbert: a universal deep-learning model of protein sequence and function.Bioinformatics, 38(8):2102–2110, 2022
Nadav Brandes, Dan Ofer, Yam Peleg, Nadav Rappoport, and Michal Linial. Proteinbert: a universal deep-learning model of protein sequence and function.Bioinformatics, 38(8):2102–2110, 2022
2022
-
[6]
This looks like that: deep learning for interpretable image recognition.Advances in neural information processing systems, 32, 2019
Chaofan Chen, Oscar Li, Daniel Tao, Alina Barnett, Cynthia Rudin, and Jonathan K Su. This looks like that: deep learning for interpretable image recognition.Advances in neural information processing systems, 32, 2019
2019
-
[7]
Structural and kinetic basis for heightened immunogenicity of t cell vaccines.The Journal of experimental medicine, 201(8):1243–1255, 2005
Ji-Li Chen, Guillaume Stewart-Jones, Giovanna Bossi, Nikolai M Lissin, Linda Wooldridge, Ed Man Lik Choi, Gerhard Held, P Rod Dunbar, Robert M Esnouf, Malkit Sami, et al. Structural and kinetic basis for heightened immunogenicity of t cell vaccines.The Journal of experimental medicine, 201(8):1243–1255, 2005
2005
-
[8]
Deep learning predictions of tcr-epitope interactions reveal epitope-specific chains in dual alpha t cells.Nature Communications, 15(1):3211, 2024
Giancarlo Croce, Sara Bobisse, Dana Léa Moreno, Julien Schmidt, Philippe Guillame, Alexandre Harari, and David Gfeller. Deep learning predictions of tcr-epitope interactions reveal epitope-specific chains in dual alpha t cells.Nature Communications, 15(1):3211, 2024
2024
-
[9]
Quantifiable predictive features define epitope-specific t cell receptor repertoires.Nature, 547(7661):89–93, 2017
Pradyot Dash, Andrew J Fiore-Gartland, Tomer Hertz, George C Wang, Shalini Sharma, Aisha Souquette, Jeremy Chase Crawford, E Bridie Clemens, Thi HO Nguyen, Katherine Kedzierska, et al. Quantifiable predictive features define epitope-specific t cell receptor repertoires.Nature, 547(7661):89–93, 2017
2017
-
[10]
T-cell antigen receptor genes and t-cell recognition.Nature, 334(6181):395– 402, 1988
Mark M Davis and Pamela J Bjorkman. T-cell antigen receptor genes and t-cell recognition.Nature, 334(6181):395– 402, 1988
1988
-
[11]
Nettcr-struc, a structure driven approach for prediction of tcr-pmhc interactions.Frontiers in Immunology, 16:1616328, 2025
Sebastian Nymann Deleuran and Morten Nielsen. Nettcr-struc, a structure driven approach for prediction of tcr-pmhc interactions.Frontiers in Immunology, 16:1616328, 2025
2025
-
[12]
Sliding-attention transformer neural architecture for predicting t cell receptor–antigen–human leucocyte antigen binding.Nature Machine Intelligence, 6(10):1216–1230, 2024
Ziyan Feng, Jingyang Chen, Youlong Hai, Xuelian Pang, Kun Zheng, Chenglong Xie, Xiujuan Zhang, Shengqing Li, Chengjuan Zhang, Kangdong Liu, et al. Sliding-attention transformer neural architecture for predicting t cell receptor–antigen–human leucocyte antigen binding.Nature Machine Intelligence, 6(10):1216–1230, 2024
2024
-
[13]
Attentive variational information bottleneck for tcr–peptide interaction prediction.Bioinformatics, 39(1):btac820, 2023
Filippo Grazioli, Pierre Machart, Anja Mösch, Kai Li, Leonardo V Castorina, Nico Pfeifer, and Martin Ren- qiang Min. Attentive variational information bottleneck for tcr–peptide interaction prediction.Bioinformatics, 39(1):btac820, 2023
2023
-
[14]
Analyzing the mycobacterium tuberculosis immune response by t-cell receptor clustering with gliph2 and genome-wide antigen screening.Nature biotechnology, 38(10):1194–1202, 2020
Huang Huang, Chunlin Wang, Florian Rubelt, Thomas J Scriba, and Mark M Davis. Analyzing the mycobacterium tuberculosis immune response by t-cell receptor clustering with gliph2 and genome-wide antigen screening.Nature biotechnology, 38(10):1194–1202, 2020
2020
-
[15]
Can we predict t cell specificity with digital biology and machine learning?Nature Reviews Immunology, 23(8):511–521, 2023
Dan Hudson, Ricardo A Fernandes, Mark Basham, Graham Ogg, and Hashem Koohy. Can we predict t cell specificity with digital biology and machine learning?Nature Reviews Immunology, 23(8):511–521, 2023
2023
-
[16]
A comparison of clustering models for inference of t cell receptor antigen specificity.ImmunoInformatics, 13:100033, 2024
Dan Hudson, Alex Lubbock, Mark Basham, and Hashem Koohy. A comparison of clustering models for inference of t cell receptor antigen specificity.ImmunoInformatics, 13:100033, 2024
2024
-
[17]
Mathias Fynbo Jensen and Morten Nielsen. Nettcr 2.2-improved tcr specificity predictions by combining pan-and peptide-specific training strategies, loss-scaling and integration of sequence similarity.bioRxiv, pages 2023–10, 2023. 9
2023
-
[18]
T cell antigen discovery.Nature methods, 18(8):873–880, 2021
Alok V Joglekar and Guideng Li. T cell antigen discovery.Nature methods, 18(8):873–880, 2021
2021
-
[19]
Tcrconv: predicting recognition between t cell receptors and epitopes using contextualized motifs.Bioinformatics, 39(1):btac788, 2023
Emmi Jokinen, Alexandru Dumitrescu, Jani Huuhtanen, Vladimir Gligorijevi´c, Satu Mustjoki, Richard Bonneau, Markus Heinonen, and Harri Lähdesmäki. Tcrconv: predicting recognition between t cell receptors and epitopes using contextualized motifs.Bioinformatics, 39(1):btac788, 2023
2023
-
[20]
Stcrdab: the structural t-cell receptor database.Nucleic acids research, 46(D1):D406–D412, 2018
Jinwoo Leem, Saulo H P de Oliveira, Konrad Krawczyk, and Charlotte M Deane. Stcrdab: the structural t-cell receptor database.Nucleic acids research, 46(D1):D406–D412, 2018
2018
-
[21]
Rational multi-modal transformers for tcr-pmhc prediction
Jiarui Li, Zixiang Yin, Zhengming Ding, Samuel J Landry, and Ramgopal R Mettu. Rational multi-modal transformers for tcr-pmhc prediction. InProceedings of the 16th ACM International Conference on Bioinformatics, Computational Biology and Health Informatics, pages 1–10, 2025
2025
-
[22]
Jiarui Li, Zixiang Yin, Haley Smith, Zhengming Ding, Samuel J Landry, and Ramgopal R Mettu. Quantifying cross-attention interaction in transformers for interpreting tcr-pmhc binding.arXiv preprint arXiv:2507.03197, 2025
-
[23]
Tcr3d 2.0: expanding the t cell receptor structure database with new structures, tools and interactions.Nucleic Acids Research, 53(D1):D604–D608, 2025
Valerie Lin, Melyssa Cheung, Ragul Gowthaman, Maya Eisenberg, Brian M Baker, and Brian G Pierce. Tcr3d 2.0: expanding the t cell receptor structure database with new structures, tools and interactions.Nucleic Acids Research, 53(D1):D604–D608, 2025
2025
-
[24]
Evolutionary-scale prediction of atomic-level protein structure with a language model.Science, 379(6637):1123–1130, 2023
Zeming Lin, Halil Akin, Roshan Rao, Brian Hie, Zhongkai Zhu, Wenting Lu, Nikita Smetanin, Robert Verkuil, Ori Kabeli, Yaniv Shmueli, et al. Evolutionary-scale prediction of atomic-level protein structure with a language model.Science, 379(6637):1123–1130, 2023
2023
-
[25]
Deep learning-based prediction of the t cell receptor–antigen binding specificity
Tianshi Lu, Ze Zhang, James Zhu, Yunguan Wang, Peixin Jiang, Xue Xiao, Chantale Bernatchez, John V Heymach, Don L Gibbons, Jun Wang, et al. Deep learning-based prediction of the t cell receptor–antigen binding specificity. Nature machine intelligence, 3(10):864–875, 2021
2021
-
[26]
Assessment of computational methods in predicting tcr–epitope binding recognition.Nature Methods, 23(1):248–259, 2026
Yanping Lu, Yuyan Wang, Meng Xu, Bingbing Xie, Yumeng Yang, Haodong Xu, and Shengbao Suo. Assessment of computational methods in predicting tcr–epitope binding recognition.Nature Methods, 23(1):248–259, 2026
2026
-
[27]
Netmhc-3.0: accurate web accessible predictions of human, mouse and monkey mhc class i affinities for peptides of length 8–11.Nucleic acids research, 36(suppl_2):W509–W512, 2008
Claus Lundegaard, Kasper Lamberth, Mikkel Harndahl, Søren Buus, Ole Lund, and Morten Nielsen. Netmhc-3.0: accurate web accessible predictions of human, mouse and monkey mhc class i affinities for peptides of length 8–11.Nucleic acids research, 36(suppl_2):W509–W512, 2008
2008
-
[28]
Tcr meta-clonotypes for biomarker discovery with tcrdist3 enabled identification of public, hla-restricted clusters of sars-cov-2 tcrs.Elife, 10:e68605, 2021
Koshlan Mayer-Blackwell, Stefan Schattgen, Liel Cohen-Lavi, Jeremy C Crawford, Aisha Souquette, Jessica A Gaevert, Tomer Hertz, Paul G Thomas, Philip Bradley, and Andrew Fiore-Gartland. Tcr meta-clonotypes for biomarker discovery with tcrdist3 enabled identification of public, hla-restricted clusters of sars-cov-2 tcrs.Elife, 10:e68605, 2021
2021
-
[29]
Barthelemy Meynard-Piganeau, Christoph Feinauer, Martin Weigt, Aleksandra M Walczak, and Thierry Mora. Tulip: A transformer-based unsupervised language model for interacting peptides and t cell receptors that generalizes to unseen epitopes.Proceedings of the National Academy of Sciences, 121(24):e2316401121, 2024
2024
-
[30]
Pip-net: Patch-based intuitive prototypes for interpretable image classification
Meike Nauta, Jörg Schlötterer, Maurice Van Keulen, and Christin Seifert. Pip-net: Patch-based intuitive prototypes for interpretable image classification. InProceedings of the IEEE/CVF Conference on Computer Vision and Pattern Recognition, pages 2744–2753, 2023
2023
-
[31]
Towards a systems understanding of mhc class i and mhc class ii antigen presentation.Nature reviews immunology, 11(12):823–836, 2011
Jacques Neefjes, Marlieke LM Jongsma, Petra Paul, and Oddmund Bakke. Towards a systems understanding of mhc class i and mhc class ii antigen presentation.Nature reviews immunology, 11(12):823–836, 2011
2011
-
[32]
Immunoinformatics: predicting peptide–mhc binding.Annual review of biomedical data science, 3(1):191–215, 2020
Morten Nielsen, Massimo Andreatta, Bjoern Peters, and Søren Buus. Immunoinformatics: predicting peptide–mhc binding.Annual review of biomedical data science, 3(1):191–215, 2020
2020
-
[33]
Lessons learned from the immrep23 tcr-epitope prediction challenge.ImmunoInformatics, 16:100045, 2024
Morten Nielsen, Anne Eugster, Mathias Fynbo Jensen, Manisha Goel, Andreas Tiffeau-Mayer, Aurelien Pelissier, Sebastiaan Valkiers, María Rodríguez Martínez, Barthélémy Meynard-Piganeeau, Victor Greiff, et al. Lessons learned from the immrep23 tcr-epitope prediction challenge.ImmunoInformatics, 16:100045, 2024
2024
-
[34]
T cell epitope predictions.Annual Review of Immunology, 38(1):123–145, 2020
Bjoern Peters, Morten Nielsen, and Alessandro Sette. T cell epitope predictions.Annual Review of Immunology, 38(1):123–145, 2020. 10
2020
-
[35]
epitcr: a highly sensitive predictor for tcr–peptide binding.Bioinformatics, 39(5):btad284, 2023
My-Diem Nguyen Pham, Thanh-Nhan Nguyen, Le Son Tran, Que-Tran Bui Nguyen, Thien-Phuc Hoang Nguyen, Thi Mong Quynh Pham, Hoai-Nghia Nguyen, Hoa Giang, Minh-Duy Phan, and Vy Nguyen. epitcr: a highly sensitive predictor for tcr–peptide binding.Bioinformatics, 39(5):btad284, 2023
2023
-
[36]
Tcr-like cars and tcr-cars targeting neoepitopes: an emerging potential.Cancer gene therapy, 28(6):581–589, 2021
Mansour Poorebrahim, Niloufar Mohammadkhani, Reza Mahmoudi, Monireh Gholizadeh, Elham Fakhr, and Angel Cid-Arregui. Tcr-like cars and tcr-cars targeting neoepitopes: an emerging potential.Cancer gene therapy, 28(6):581–589, 2021
2021
-
[37]
Alexander Rives, Joshua Meier, Tom Sercu, Siddharth Goyal, Zeming Lin, Jason Liu, Demi Guo, Myle Ott, C Lawrence Zitnick, Jerry Ma, et al. Biological structure and function emerge from scaling unsupervised learning to 250 million protein sequences.Proceedings of the National Academy of Sciences, 118(15):e2016239118, 2021
2021
-
[38]
Personalized rna neoantigen vaccines stimulate t cells in pancreatic cancer.Nature, 618(7963):144–150, 2023
Luis A Rojas, Zachary Sethna, Kevin C Soares, Cristina Olcese, Nan Pang, Erin Patterson, Jayon Lihm, Nicholas Ceglia, Pablo Guasp, Alexander Chu, et al. Personalized rna neoantigen vaccines stimulate t cells in pancreatic cancer.Nature, 618(7963):144–150, 2023
2023
-
[39]
Stop explaining black box machine learning models for high stakes decisions and use interpretable models instead.Nature machine intelligence, 1(5):206–215, 2019
Cynthia Rudin. Stop explaining black box machine learning models for high stakes decisions and use interpretable models instead.Nature machine intelligence, 1(5):206–215, 2019
2019
-
[40]
Unpaired tcrα+ tcrβ sequencing is sufficient for training machine learning tcr-epitope recognition predictors.bioRxiv, pages 2026–03, 2026
Aisha Shah, Raphael Genolet, Aymeric Auger, Dana Léa Moreno, Yan Liu, Giancarlo Croce, Julien Racle, Alexandre Harari, and David Gfeller. Unpaired tcrα+ tcrβ sequencing is sufficient for training machine learning tcr-epitope recognition predictors.bioRxiv, pages 2026–03, 2026
2026
-
[41]
Stag-llm: Predicting tcr-phla binding with protein language models and computationally generated 3d structures.Computational and Structural Biotechnology Journal, 2025
Jared K Slone, Minying Zhang, Peixin Jiang, Amanda Montoya, Emily Bontekoe, Barbara Nassif Rausseo, Alexandre Reuben, and Lydia E Kavraki. Stag-llm: Predicting tcr-phla binding with protein language models and computationally generated 3d structures.Computational and Structural Biotechnology Journal, 2025
2025
-
[42]
Contribution of t cell receptor alpha and beta cdr3, mhc typing, v and j genes to peptide binding prediction.Frontiers in immunology, 12:664514, 2021
Ido Springer, Nili Tickotsky, and Yoram Louzoun. Contribution of t cell receptor alpha and beta cdr3, mhc typing, v and j genes to peptide binding prediction.Frontiers in immunology, 12:664514, 2021
2021
-
[43]
Mcpas-tcr: a manually curated catalogue of pathology-associated t cell receptor sequences.Bioinformatics, 33(18):2924–2929, 2017
Nili Tickotsky, Tal Sagiv, Jaime Prilusky, Eric Shifrut, and Nir Friedman. Mcpas-tcr: a manually curated catalogue of pathology-associated t cell receptor sequences.Bioinformatics, 33(18):2924–2929, 2017
2017
-
[44]
The immune epitope database (iedb): 2018 update
Randi Vita, Swapnil Mahajan, James A Overton, Sandeep Kumar Dhanda, Sheridan Martini, Jason R Cantrell, Daniel K Wheeler, Alessandro Sette, and Bjoern Peters. The immune epitope database (iedb): 2018 update. Nucleic acids research, 47(D1):D339–D343, 2019
2018
-
[45]
Fast and accurate modeling of tcr-peptide-mhc complexes using tfold-tcr.bioRxiv, pages 2025–01, 2025
Fandi Wu, Yu Zhao, Yang Xiao, Chenchen Qin, Fang Wang, Zihan Wu, Long-Kai Huang, Xiao Liu, Jiangning Song, Bing He, et al. Fast and accurate modeling of tcr-peptide-mhc complexes using tfold-tcr.bioRxiv, pages 2025–01, 2025
2025
-
[46]
Tcrmodel2: high-resolution modeling of t cell receptor recognition using deep learning.Nucleic Acids Research, 51(W1):W569– W576, 2023
Rui Yin, Helder V Ribeiro-Filho, Valerie Lin, Ragul Gowthaman, Melyssa Cheung, and Brian G Pierce. Tcrmodel2: high-resolution modeling of t cell receptor recognition using deep learning.Nucleic Acids Research, 51(W1):W569– W576, 2023
2023
-
[47]
Giana allows computationally-efficient tcr clustering and multi-disease repertoire classification by isometric transformation.Nature communications, 12(1):4699, 2021
Hongyi Zhang, Xiaowei Zhan, and Bo Li. Giana allows computationally-efficient tcr clustering and multi-disease repertoire classification by isometric transformation.Nature communications, 12(1):4699, 2021
2021
-
[48]
cannot find core
Wei Zhang, Longlong Wang, Ke Liu, Xiaofeng Wei, Kai Yang, Wensi Du, Shiyu Wang, Nannan Guo, Chuanchuan Ma, Lihua Luo, et al. Pird: pan immune repertoire database.Bioinformatics, 36(3):897–903, 2020. 11 Structure-Regularized Interpretable TCR-Epitope Prediction (Supplementary) Jiarui Li1, Zixiang Yin1, Yunbei Zhang1, Janet Wang1 Samuel J. Landry2, Zhengmin...
2020
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.