Active diffusion enhances plankton carbon capture and phycosphere radius
Pith reviewed 2026-06-29 01:48 UTC · model grok-4.3
The pith
Active mixing by plankton increases carbon uptake and enlarges the phycosphere radius.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
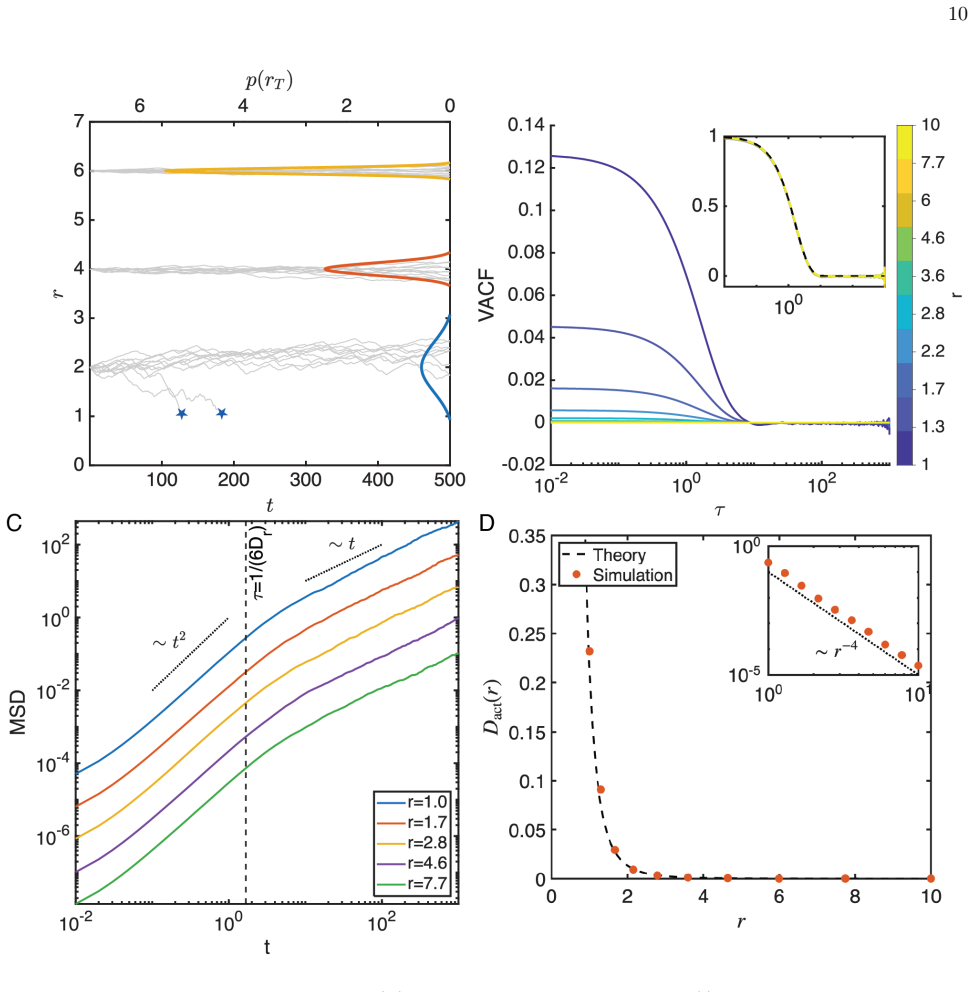
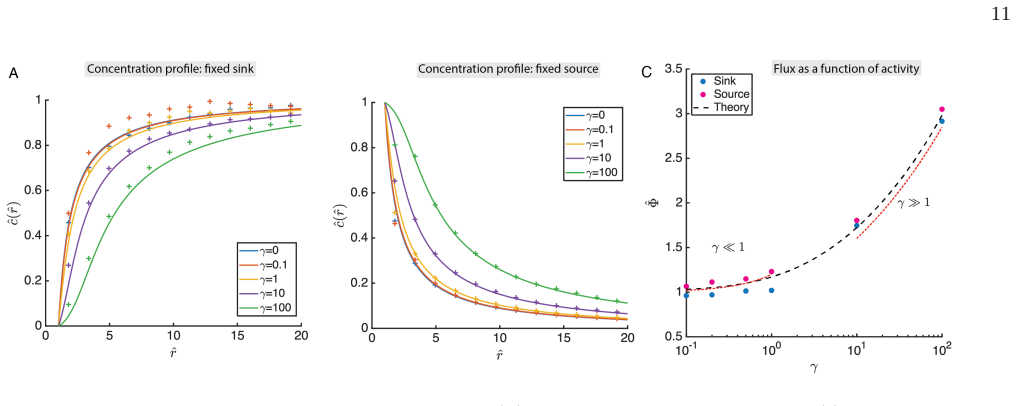
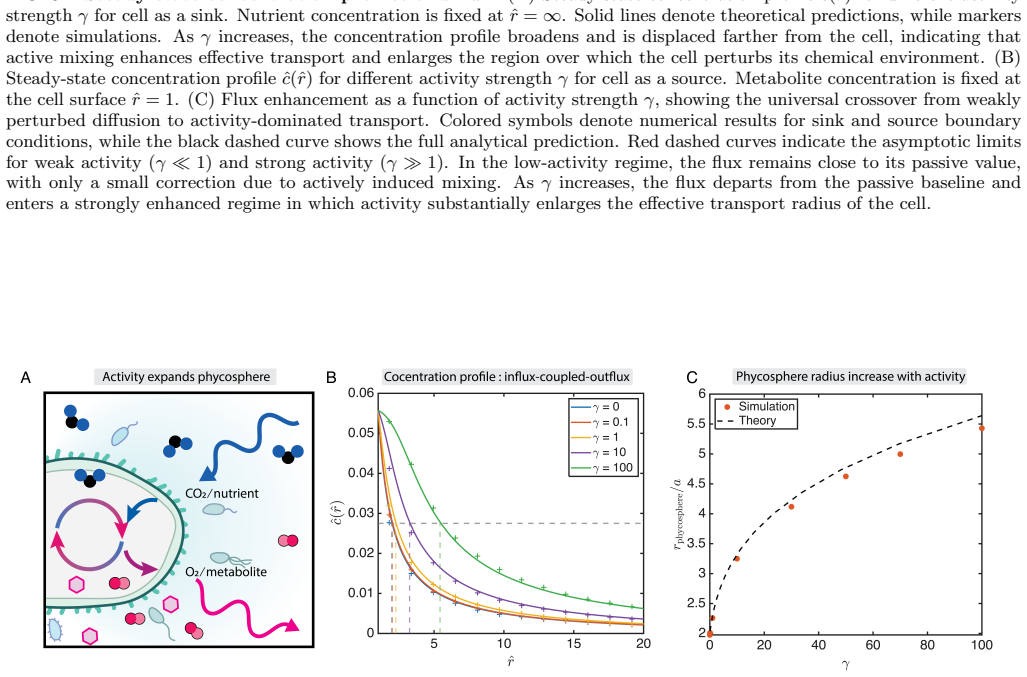
By developing exact analytical expressions validated by stochastic simulations, we determine the enhanced diffusivity of phycosphere particles as a function of mixing activity, and their resulting fluxes and concentration fields. Hence, we find that plankton can significantly increase their uptake and photosynthetic turnover. Moreover, we find that the phycosphere radius is enlarged both by increased metabolism and by increased diffusive transport further from the organism.
What carries the argument
exact analytical expressions for the enhanced diffusivity of phycosphere particles as a function of mixing activity
If this is right
- Plankton significantly increase their carbon uptake and photosynthetic turnover through active mixing.
- The phycosphere radius enlarges due to increased metabolism from higher fluxes.
- The phycosphere radius also enlarges due to increased diffusive transport further from the organism.
- These results supply new biophysical insights into marine microbial ecology.
- Active mixing carries implications for estimates of global carbon capture and climate change.
Where Pith is reading between the lines
- Ocean carbon-cycle models may need to include active mixing to avoid underestimating plankton fixation rates.
- The same analytical treatment of enhanced diffusivity could be applied to other microorganisms that exchange solutes across a boundary layer.
- Energy costs of sustained mixing may impose an upper bound on the enhancement that real cells can achieve.
Load-bearing premise
Mixing activity can be parameterized to allow exact analytical expressions for enhanced diffusivity without additional biological limits on feasible stirring rates or energy costs.
What would settle it
Direct measurements of carbon uptake rates and phycosphere radius in live plankton at controlled levels of active mixing, compared with the analytical predictions.
Figures
read the original abstract
Plankton fix about 40 gigatons of carbon annually, using photosynthesis to convert $\text{CO}_2$ into $\text{O}_2$ and carbohydrates. These solutes are exchanged with the ocean in a diffusive boundary layer around the organism called the phycosphere. Here, we study how organisms can increase their carbon influx and outflux by actively mixing the surrounding fluid. By developing exact analytical expressions validated by stochastic simulations, we determine the enhanced diffusivity of phycosphere particles as a function of mixing activity, and their resulting fluxes and concentration fields. Hence, we find that plankton can significantly increase their uptake and photosynthetic turnover. Moreover, we find that the phycosphere radius is enlarged both by increased metabolism and by increased diffusive transport further from the organism. These results provide new biophysical insights into marine microbial ecology, with important implications for global carbon capture and climate change.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops exact analytical expressions for the enhanced diffusivity of particles in the phycosphere as a function of a mixing activity parameter, validates them via stochastic simulations, and concludes that active mixing allows plankton to significantly increase carbon uptake, photosynthetic turnover, and phycosphere radius through both metabolic and transport effects.
Significance. If the derivations are rigorous and the mixing activity levels are biologically attainable, the work supplies new biophysical mechanisms linking organism-scale fluid mixing to carbon fluxes, with potential consequences for models of marine microbial ecology and global carbon cycling. The attempt at closed-form expressions plus simulation cross-check is a methodological strength.
major comments (3)
- [Abstract / mixing parameterization] Abstract and the section defining the mixing activity: the enhanced-diffusivity expressions treat mixing activity as an unbounded free parameter whose only limit is mathematical convergence. No metabolic power budget, flagellar energy cost, or cell-scale Reynolds-number constraint is imposed, so the reported flux gains may not be attainable by real plankton; this is load-bearing for the central claim that active mixing 'significantly increases' uptake.
- [Results on fluxes and phycosphere radius] Results on flux and radius: the enlargement of the phycosphere is attributed to both increased metabolism and increased diffusive transport, yet the mixing activity itself is not shown to be independently measurable or constrained; if it is instead chosen to match observed fluxes, the 'enhanced diffusivity' reduces to a fitted quantity by construction, undermining the predictive claim.
- [Simulation validation] Simulation validation section: the abstract asserts that stochastic simulations confirm the exact analytical expressions, but without reported parameter ranges, convergence checks, or explicit comparison metrics it is impossible to assess whether the validation is independent or post-hoc.
minor comments (1)
- [Methods] Notation for the mixing activity parameter should be introduced with a clear symbol and units in the first equation where it appears.
Simulated Author's Rebuttal
We thank the referee for the constructive feedback. We address each major comment below and will revise the manuscript accordingly to improve clarity, add necessary details, and strengthen the presentation of biological constraints.
read point-by-point responses
-
Referee: [Abstract / mixing parameterization] Abstract and the section defining the mixing activity: the enhanced-diffusivity expressions treat mixing activity as an unbounded free parameter whose only limit is mathematical convergence. No metabolic power budget, flagellar energy cost, or cell-scale Reynolds-number constraint is imposed, so the reported flux gains may not be attainable by real plankton; this is load-bearing for the central claim that active mixing 'significantly increases' uptake.
Authors: We agree that the mixing activity parameter requires biological grounding to support claims of significant increases. The current work is a theoretical derivation of the functional dependence; in revision we will add a dedicated subsection with literature-based estimates of realistic mixing activity values drawn from flagellar propulsion studies, including order-of-magnitude power budgets and Reynolds-number limits for representative plankton. This will explicitly bound the parameter and show which flux gains remain attainable. revision: yes
-
Referee: [Results on fluxes and phycosphere radius] Results on flux and radius: the enlargement of the phycosphere is attributed to both increased metabolism and increased diffusive transport, yet the mixing activity itself is not shown to be independently measurable or constrained; if it is instead chosen to match observed fluxes, the 'enhanced diffusivity' reduces to a fitted quantity by construction, undermining the predictive claim.
Authors: The mixing activity parameter is defined as an independent input (quantifying the strength of active fluid stirring) that can be measured separately, for example via particle-image velocimetry of the flow field around the cell. The analytical expressions then predict the resulting diffusivity, fluxes, and phycosphere radius; no fitting to observed carbon fluxes is performed. We will add explicit language and an example calculation clarifying the direction of prediction and how activity could be constrained independently of flux measurements. revision: partial
-
Referee: [Simulation validation] Simulation validation section: the abstract asserts that stochastic simulations confirm the exact analytical expressions, but without reported parameter ranges, convergence checks, or explicit comparison metrics it is impossible to assess whether the validation is independent or post-hoc.
Authors: We accept that the validation section lacks sufficient technical detail for independent assessment. In the revised manuscript we will expand this section to report the full range of mixing activity values simulated, the time-step and ensemble sizes used, convergence criteria (e.g., steady-state tolerance on concentration profiles), and quantitative comparison metrics such as maximum relative error and L2-norm difference between the analytical and simulated enhanced diffusivities. revision: yes
Circularity Check
No circularity: analytical model treats mixing activity as independent input parameter
full rationale
The paper derives exact analytical expressions for enhanced diffusivity D_eff as a function of a mixing activity parameter, then computes resulting fluxes and phycosphere radius. These expressions are validated against stochastic simulations rather than fitted to the target carbon-uptake data. No self-citation chain, no uniqueness theorem imported from prior work, and no renaming of known results as new derivations. The mixing activity remains a free parameter whose biological feasibility is outside the derivation; the mathematical steps do not reduce the output to the input by construction. This is a standard forward-modeling approach with independent content.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
Fixed surface concentration The same transport framework can also be used to describe the dispersion of a scalar released from the cell surface. As a complementary problem, we consider a fixed-concentration source, with boundary conditions c(a) =C a, c(∞) = 0,corresponding to a cell surface maintained at concentrationC ∞ and a vanishing far-field concentr...
-
[2]
Let the outward flux density at the cell bew
Constant surface flux Instead of prescribing the concentration at the cell sur- face, another model for metabolic waste release is to pre- 6 scribe the secretion flux. Let the outward flux density at the cell bew. The total secretion rate is Φ out = 4πa2w. Solving the same radial diffusion equation with bound- ary conditions−D(a) dc dr r=a =w, c(∞) = 0.gi...
-
[3]
G. Hays, A. Richardson, and C. Robinson, Climate change and marine plankton, Trends in Ecology & Evo- lution20, 337 (2005)
2005
-
[4]
J. S. Guasto, R. Rusconi, and R. Stocker, Fluid Me- chanics of Planktonic Microorganisms, Annual Review of Fluid Mechanics44, 373 (2012)
2012
-
[5]
Rusconi and R
R. Rusconi and R. Stocker, Microbes in flow, Current Opinion in Microbiology25, 1 (2015)
2015
-
[6]
J. D. Wheeler, E. Secchi, R. Rusconi, and R. Stocker, Not Just Going with the Flow: The Effects of Fluid Flow on Bacteria and Plankton, Annual Review of Cell and Developmental Biology35, 213 (2019). 7
2019
-
[7]
F. Azam, T. Fenchel, J. G. Field, J. S. Gray, L. A. Meyer- Reil, and F. Thingstad, The ecological role of water- column microbes in the sea, Marine Ecology Progress Series10, 257 (1983)
1983
-
[8]
Buchan, G
A. Buchan, G. R. LeCleir, C. A. Gulvik, and J. M. Gon- zalez, Master recyclers: features and functions of bacteria associated with phytoplankton blooms, Nature Reviews Microbiology12, 686 (2014)
2014
-
[9]
N. Jiao, T. Luo, Q. Chen, Z. Zhao, X. Xiao, J. Liu, Z. Jian, S. Xie, H. Thomas, G. J. Herndl, R. Benner, M. Gonsior, F. Chen, W.-J. Cai, and C. Robinson, The microbial carbon pump and climate change, Nature Re- views Microbiology22, 408 (2024)
2024
-
[10]
J. R. Seymour, S. A. Amin, J. B. Raina, and R. Stocker, Zooming in on the phycosphere: The ecological interface for phytoplankton-bacteria relationships (2017)
2017
-
[11]
Smriga, V
S. Smriga, V. I. Fernandez, J. G. Mitchell, and R. Stocker, Chemotaxis toward phytoplankton drives or- ganic matter partitioning among marine bacteria, Pro- ceedings of the National Academy of Sciences of the United States of America113, 1576 (2016)
2016
-
[12]
S. A. Amin, L. R. Hmelo, H. M. van Tol, B. P. Durham, L. T. Carlson, K. R. Heal, R. L. Morales, C. T. Berthi- aume, M. S. Parker, B. Djunaedi, A. E. Ingalls, M. R. Parsek, M. A. Moran, and E. V. Armbrust, Interaction and signalling between a cosmopolitan phytoplankton and associated bacteria, Nature522, 98 (2015)
2015
-
[13]
J. C. Prairie, K. R. Sutherland, K. J. Nickols, and A. M. Kaltenberg, Biophysical interactions in the plankton: A cross-scale review, Limnology and Oceanography: Fluids and Environments2, 121 (2012)
2012
-
[14]
Stocker, Marine microbes see a sea of gradients, Sci- ence338, 628 (2012)
R. Stocker, Marine microbes see a sea of gradients, Sci- ence338, 628 (2012)
2012
-
[15]
Stocker, J
R. Stocker, J. R. Seymour, A. Samadani, D. E. Hunt, and M. F. Polz,Rapid chemotactic response enables marine bacteria to exploit ephemeral microscale nutrient patches, Tech. Rep. (2008)
2008
-
[16]
Forget, O
M. Forget, O. M¨ uller, and R. Stocker, The ecology of bacterial attachment to phytoplankton, Nature Microbi- ology11, 1180 (2026)
2026
-
[17]
Dow, How do quorum-sensing signals mediate algae– bacteria interactions? (2021)
L. Dow, How do quorum-sensing signals mediate algae– bacteria interactions? (2021)
2021
-
[18]
Semple and J
A. Semple and J. Pandhal, Engineering the phycosphere: fundamental concepts and tools for the bottom-up design of microalgal-bacterial consortia, Applied Phycology6, 21 (2025)
2025
-
[19]
Passow,Progress in Oceanography, Tech
U. Passow,Progress in Oceanography, Tech. Rep. (2002)
2002
-
[20]
A. W. Decho and T. Gutierrez, Microbial Extracel- lular Polymeric Substances (EPSs) in Ocean Systems, Frontiers in Microbiology8, 10.3389/fmicb.2017.00922 (2017)
-
[21]
Kiørboe and G
T. Kiørboe and G. A. Jackson, Marine snow, organic so- lute plumes, and optimal chemosensory behavior of bac- teria, Limnology and Oceanography46, 1309 (2001)
2001
-
[22]
P. W. Boyd, H. Claustre, M. Levy, D. A. Siegel, and T. Weber, Multi-faceted particle pumps drive carbon se- questration in the ocean, Nature568, 327 (2019)
2019
-
[23]
A. L. Alldredge and M. W. Silver, Characteristics, dy- namics and significance of marine snow, Progress in Oceanography20, 41 (1988)
1988
-
[24]
Azam and R
F. Azam and R. A. Long, Sea snow microcosms, Nature 414, 495 (2001)
2001
-
[25]
Simon, H
M. Simon, H. Grossart, B. Schweitzer, and H. Ploug, Mi- crobial ecology of organic aggregates in aquatic ecosys- tems, Aquatic Microbial Ecology28, 175 (2002)
2002
-
[26]
D. C. Smith, M. Simon, A. L. Alldredge, and F. Azam, Intense hydrolytic enzyme activity on marine aggregates and implications for rapid particle dissolution, Nature 359, 139 (1992)
1992
-
[27]
H. C. Berg and E. M. Purcell, Physics of chemoreception, Biophysical Journal20, 193 (1977)
1977
-
[28]
W. H. Munk and G. A. Riley, Absorption of nutrients by aquatic plants, Journal of Marine Research11, 215 (1952)
1952
-
[29]
W. J. Pasciak and J. Gavis, Transport limitation of nu- trient uptake in phytoplankton, Limnology and Oceanog- raphy19, 881 (1974)
1974
-
[30]
Karp-Boss, E
L. Karp-Boss, E. Boss, and P. A. Jumars, Nutrient fluxes to planktonic osmotrophs in the presence of fluid motion, Oceanography and Marine Biology: An Annual Review 34, 71 (1996)
1996
-
[31]
C. Lindemann, Ø. Fiksen, K. H. Andersen, and D. L. Aksnes, Scaling laws in phytoplankton nutri- ent uptake affinity, Frontiers in Marine Science3, 10.3389/fmars.2016.00026 (2016)
-
[32]
Gavis,Munk and Riley revisited: nutrient diffusion transport and rates of phytoplankton growth, Tech
J. Gavis,Munk and Riley revisited: nutrient diffusion transport and rates of phytoplankton growth, Tech. Rep. (1976)
1976
-
[33]
M. N. Breckels, D. E. Boakes, E. A. Codling, G. Ma- lin, S. D. Archer, and M. Steinke, Modelling the concen- tration of exuded dimethylsulphoniopropionate (DMSP) in the boundary layer surrounding phytoplankton cells, Journal of Plankton Research32, 253 (2010)
2010
-
[34]
R. E. Goldstein, Green algae as model organisms for bi- ological fluid dynamics (2015)
2015
-
[35]
Hartmann, ¨O
C. Hartmann, ¨O. ¨Ozmutlu, H. Petermeier, J. Fried, and A. Delgado, Analysis of the flow field induced by the ses- sile peritrichous ciliate Opercularia asymmetrica, Journal of Biomechanics40, 137 (2007)
2007
-
[36]
Magar, T
V. Magar, T. Goto, and T. J. Pedley, Nutrient uptake by a self-propelled steady squirmer, Quarterly Journal of Mechanics and Applied Mathematics56, 65 (2003)
2003
-
[37]
J. Liu, Y. Man, J. H. Costello, and E. Kanso, Feeding Rates in Sessile versus Motile Ciliates are Hydrodynam- ically Equivalent (2024)
2024
-
[39]
Thiffeault and S
J.-L. Thiffeault and S. Childress,Stirring by swimming bodies, Tech. Rep. (2010)
2010
-
[40]
Z. Lin, J. L. Thiffeault, and S. Childress, Stirring by squirmers, Journal of Fluid Mechanics669, 167 (2011)
2011
-
[41]
I. M. Zaid, J. Dunkel, and J. M. Yeomans, L´ evy fluc- tuations and mixing in dilute suspensions of algae and bacteria (2011)
2011
-
[42]
D. O. Pushkin and J. M. Yeomans, Fluid mixing by curved trajectories of microswimmers, Physical Review Letters111, 10.1103/PhysRevLett.111.188101 (2013)
-
[43]
Kanazawa, T
K. Kanazawa, T. G. Sano, A. Cairoli, and A. Baule, Loopy L´ evy flights enhance tracer diffusion in active sus- pensions, Nature579, 364 (2020)
2020
-
[44]
C. Jin, Y. Chen, C. C. Maass, and A. J. Mathijssen, Col- lective Entrainment and Confinement Amplify Transport by Schooling Microswimmers, Physical Review Letters 127, 10.1103/PhysRevLett.127.088006 (2021). 8
- [45]
-
[46]
K. Drescher, R. E. Goldstein, N. Michel, M. Polin, and I. Tuval, Direct measurement of the flow field around swimming microorganisms, Physical Review Letters105, 10.1103/PhysRevLett.105.168101 (2010)
-
[47]
Drescher, J
K. Drescher, J. Dunkel, L. H. Cisneros, S. Ganguly, R. E. Goldstein, R. E. G. Designed, and R. E. G. Performed, Fluid dynamics and noise in bacterial cell-cell and cell- surface scattering, Proceedings of the National Academy of Sciences of the United States of America108, 10940 (2011)
2011
-
[48]
A. J. Mathijssen, F. Guzm´ an-Lastra, A. Kaiser, and H. L¨ owen, Nutrient Transport Driven by Mi- crobial Active Carpets, Physical Review Letters121, 10.1103/PhysRevLett.121.248101 (2018)
-
[49]
F. Guzm´ an-Lastra, H. L¨ owen, and A. J. Mathijssen, Active carpets drive non-equilibrium diffusion and en- hanced molecular fluxes, Nature Communications12, 10.1038/s41467-021-22029-y (2021)
-
[50]
Aguayo, A
G. Aguayo, A. Mathijssen, H. Ulloa, R. Soto, and F. Guzm´ an-Lastra, Floating active carpets drive trans- port and aggregation in aquatic ecosystems, Journal of Fluid Mechanics995, A16 (2024)
2024
-
[51]
F. A. Barros, H. N. Ulloa, G. Aguayo, A. J. Mathijssen, and F. Guzm´ an-Lastra, Layered aquatic microenviron- ments control fluctuations generated by active carpets, Physical Review Research7, 013152 (2025)
2025
-
[52]
F. A. Barros, I. Salas, E. Lushi, and F. Guzm´ an-Lastra, Confined floating active carpets generate coherent vor- tical flows that enhance transport, Journal of Physics: Condensed Matter38, 095101 (2026)
2026
-
[53]
Lauga and T
E. Lauga and T. R. Powers, The hydrodynamics of swim- ming microorganisms, Reports on Progress in Physics72, 096601 (2009)
2009
-
[54]
Elgeti, R
J. Elgeti, R. G. Winkler, and G. Gompper, Physics of microswimmers—single particle motion and collective behavior: a review, Reports on Progress in Physics78, 056601 (2015)
2015
-
[55]
Bruot and P
N. Bruot and P. Cicuta, Realizing the Physics of Motile Cilia Synchronization with Driven Colloids, Annual Re- view of Condensed Matter Physics7, 323 (2016)
2016
-
[56]
Gilpin, M
W. Gilpin, M. S. Bull, and M. Prakash, The multiscale physics of cilia and flagella, Nature Reviews Physics2, 74 (2020)
2020
-
[57]
Wadhwa and H
N. Wadhwa and H. C. Berg, Bacterial motility: machin- ery and mechanisms, Nature Reviews Microbiology20, 161 (2022)
2022
-
[58]
Lauga,The Fluid Dynamics of Cell Motility(Cam- bridge University Press, 2020)
E. Lauga,The Fluid Dynamics of Cell Motility(Cam- bridge University Press, 2020)
2020
-
[59]
B. J. Berne and R. Pecora,Dynamic light scattering : with applications to chemistry, biology, and physics(Wi- ley, 1976) p. 376
1976
-
[60]
E. Lushi, H. Wioland, and R. E. Goldstein, Fluid flows created by swimming bacteria drive self- organization in confined suspensions, Proceedings of the National Academy of Sciences111, 9733 (2014), https://www.pnas.org/doi/pdf/10.1073/pnas.1405698111
-
[61]
R. Stocker and J. R. Seymour, Ecology and physics of bacterial chemotaxis in the ocean, Microbiology and Molecular Biology Reviews76, 792 (2012), https://journals.asm.org/doi/pdf/10.1128/mmbr.00029- 12
-
[62]
Liu, Data availability (2025)
M. Liu, Data availability (2025)
2025
-
[63]
Meerschaert and H
M. Meerschaert and H. P. Scheffler,Handbook of Frac- tional Calculus with Applications, Tech. Rep
-
[64]
G. Vaccario, C. Antoine, and J. Talbot, First-Passage Times in d -Dimensional Heterogeneous Media, Physical Review Letters115, 10.1103/PhysRevLett.115.240601 (2015)
-
[65]
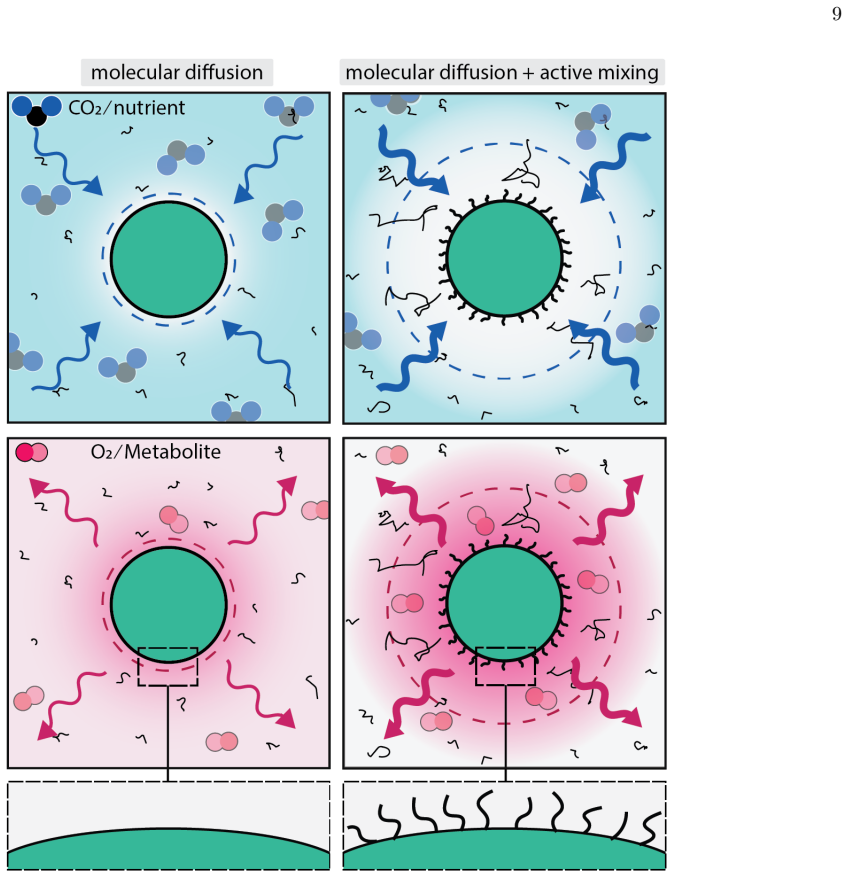
A. W. Lau and T. C. Lubensky, State-dependent dif- fusion: Thermodynamic consistency and its path in- tegral formulation, Physical Review E - Statistical, Nonlinear, and Soft Matter Physics76, 10.1103/Phys- RevE.76.011123 (2007). 9 FIG. 1.Active surface forcing creates a heterogeneous transport region.Schematic comparing purely thermal diffusion (left col...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.