A nonlinear theory for chemotactic fronts of mixed populations
Pith reviewed 2026-06-26 21:52 UTC · model grok-4.3
The pith
Heterogeneity in diffusivity, consumption, and sensitivity produces four regimes for chemotactic density profiles in mixed cell populations.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
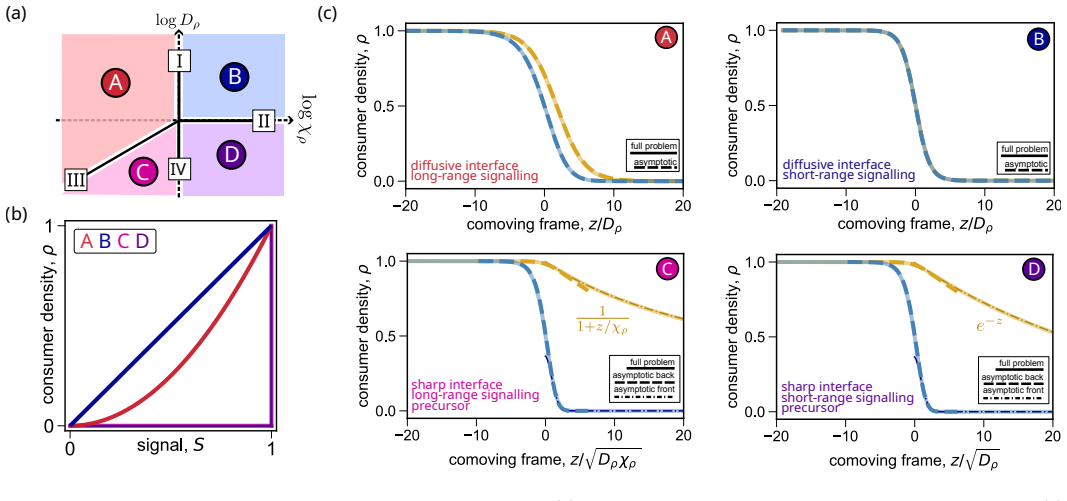
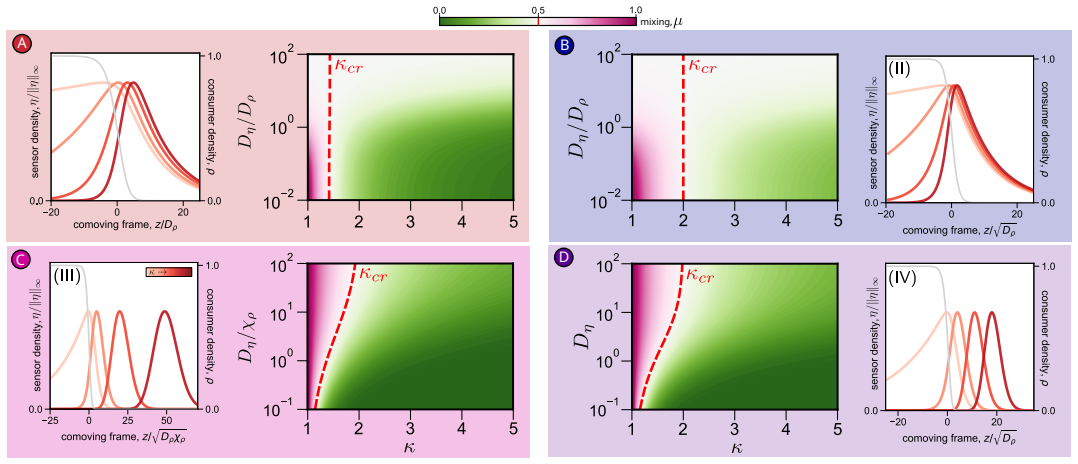
Using asymptotic analysis, the full system of PDEs for heterogeneous chemotactic migration is reduced to a closed nonlinear theory. This theory reveals four distinct dynamical behaviours determined by the relative magnitudes of heterogeneity in cell diffusivity, chemoattractant consumption, and chemotactic sensitivity, which together classify all possible regimes of density profile formation.
What carries the argument
The asymptotic reduction of the PDE system to a closed nonlinear theory that yields four dynamical regimes for the density profiles.
If this is right
- Density profiles of mixed populations can be predicted from the three heterogeneity parameters.
- The co-migration of dendritic and T cells operates in a regime balancing mixing and T-cell localization at the front.
- Strong consumption by dendritic cells generates intermediate long-range chemoattractant signaling.
- All possible collective migration patterns fall into one of the four regimes.
Where Pith is reading between the lines
- The four-regime classification could guide experiments to engineer desired spatial organizations in cell collectives.
- Similar non-reciprocal interactions may govern other biological systems involving heterogeneous populations.
- If validated, the theory suggests that tuning consumption rates can control the extent of mixing versus segregation.
Load-bearing premise
The asymptotic analysis produces a closed theory without residual higher-order effects altering the density profiles across all parameter combinations.
What would settle it
Finding a set of measured parameters for a cell collective whose observed density profile does not match any of the four predicted regime profiles.
Figures
read the original abstract
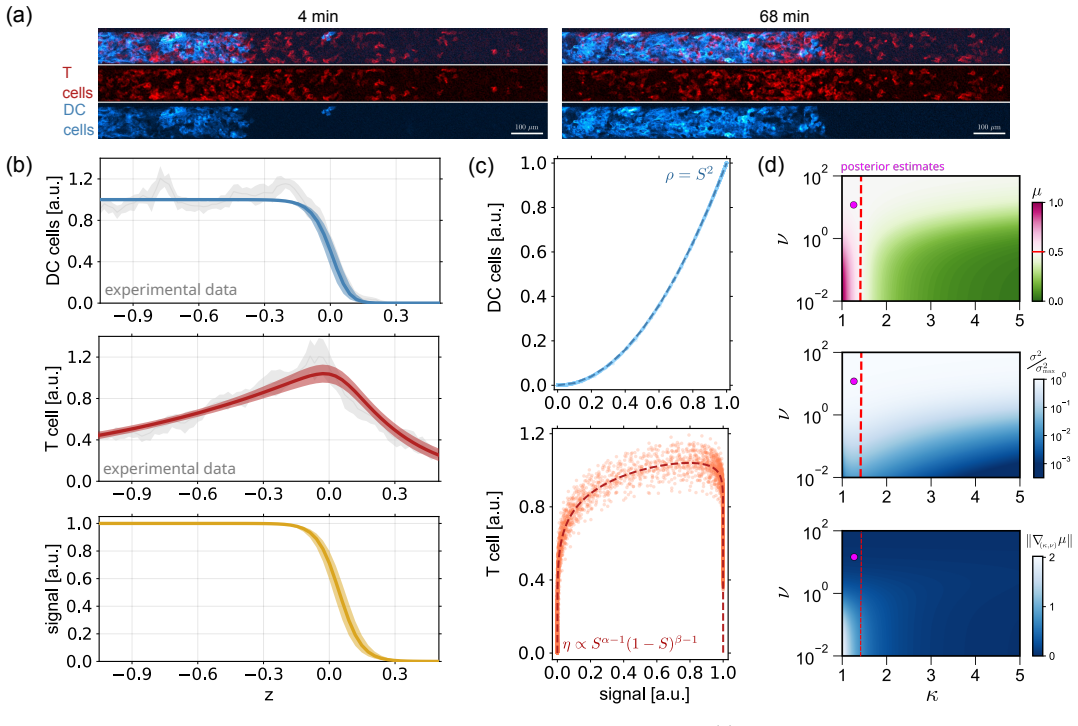
Collective migration of heterogeneous cell populations is central to many biological and physiological processes, including development and immune response. Recent experimental and theoretical advances have shown how asymmetric interactions with self-generated chemical gradients shape the spatial distribution of distinct cell types within migrating collectives. However, the principles governing robust spatial organisation of heterogeneous cell populations remain poorly understood. Here, we use asymptotic analysis to systematically derive a nonlinear analytical theory for heterogeneous cell collectives guided by self-generated chemotaxis. Our theory disentangles how heterogeneity in cell diffusivity, chemoattractant consumption, and chemotactic sensitivity shape the density profiles of migrating heterogeneous collectives, revealing four distinct dynamical behaviours that together capture all possible regimes. We calibrate our framework to experimental data on the co-migration of dendritic and T cells. We predict that this system operates in a parameter regime that balances intercellular mixing with T-cell localisation at the leading front of the migrating collective. Our theory reveals that this behaviour is enabled by intermediate long-range chemoattractant signalling generated through strong chemoattractant consumption by dendritic cells. Overall, our framework provides general principles for understanding how non-reciprocal chemical interactions shape robust collective migration in heterogeneous cell populations.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript uses asymptotic analysis to derive a nonlinear analytical theory for chemotactic fronts in heterogeneous cell populations. It identifies four distinct dynamical regimes arising from heterogeneity in cell diffusivity, chemoattractant consumption, and chemotactic sensitivity, claiming these regimes together exhaustively classify all possible parameter combinations. The theory is calibrated to experimental data on dendritic and T cell co-migration and predicts that this system operates in a regime balancing intercellular mixing with T-cell localisation at the leading front, enabled by intermediate long-range signalling from strong consumption by dendritic cells.
Significance. If the asymptotic reductions are valid and the four regimes are exhaustive without residual higher-order effects, the work supplies general principles for how non-reciprocal chemical interactions organise spatial structure in mixed collectives. The calibration to immune-cell data and the concrete prediction for the dendritic-T cell system are strengths that increase the result's applicability to developmental and immunological contexts.
major comments (2)
- [Regime classification section] The section deriving the four regimes (likely the main results section following the asymptotic analysis): the claim that these regimes 'together capture all possible regimes' requires an explicit demonstration that the chosen scalings remain uniformly valid and that no hybrid behaviours arise at regime boundaries where competing effects enter at the same asymptotic order; without this, the exhaustiveness assertion is not yet load-bearing.
- [Application to dendritic-T cell data] The calibration and prediction paragraph for the dendritic-T cell system: it is unclear whether the assigned regime follows from independently measured parameter values or from fitting consumption and sensitivity to the same density profiles used for validation; this distinction is needed to confirm the prediction is falsifiable rather than post-hoc.
minor comments (2)
- Figure captions should explicitly indicate which of the four regimes each panel corresponds to and reference the relevant reduced equations.
- All asymptotic scalings and matching conditions should be numbered and cross-referenced in the text for traceability.
Simulated Author's Rebuttal
We thank the referee for their thoughtful and constructive report. We address each major comment in detail below and have revised the manuscript to strengthen the presentation of the regime classification and the calibration procedure.
read point-by-point responses
-
Referee: [Regime classification section] The section deriving the four regimes (likely the main results section following the asymptotic analysis): the claim that these regimes 'together capture all possible regimes' requires an explicit demonstration that the chosen scalings remain uniformly valid and that no hybrid behaviours arise at regime boundaries where competing effects enter at the same asymptotic order; without this, the exhaustiveness assertion is not yet load-bearing.
Authors: We agree that an explicit check of uniform validity and boundary behaviour is required to make the exhaustiveness claim rigorous. In the revised manuscript we have added a new subsection (Section 3.5) together with Supplementary Note S4. There we perform a matched-asymptotic analysis at each regime boundary, retaining the next-order terms when two effects become comparable. This shows that the transition between regimes occurs smoothly at leading order and that no additional hybrid scalings appear; the four regimes therefore remain exhaustive. The added material is referenced from the main-text claim. revision: yes
-
Referee: [Application to dendritic-T cell data] The calibration and prediction paragraph for the dendritic-T cell system: it is unclear whether the assigned regime follows from independently measured parameter values or from fitting consumption and sensitivity to the same density profiles used for validation; this distinction is needed to confirm the prediction is falsifiable rather than post-hoc.
Authors: We have revised the relevant paragraph (now Section 4.2) to state explicitly that the consumption rates and chemotactic sensitivities are taken from independent literature values measured in separate assays (cited references), while the co-migration density profiles serve only for post-prediction validation. The regime assignment is therefore a genuine prediction based on independently determined parameter orderings. We have also added a short discussion of how future experiments could falsify the predicted regime. revision: yes
Circularity Check
Asymptotic reduction from PDEs to closed nonlinear theory is independent of target regimes and data calibration
full rationale
The derivation begins from a system of PDEs for cell densities and chemoattractant, applies asymptotic analysis with stated scalings on diffusivity, consumption, and sensitivity parameters, and obtains four reduced regimes. This reduction is performed once from the governing equations and does not invoke the experimental data or the specific dendritic-T cell prediction. Calibration occurs after the theory is derived and is used only to assign parameter values; the claim that the four regimes exhaust all combinations follows from the asymptotic balances, not from fitting. No self-citation chain, self-definitional step, or fitted quantity renamed as prediction is present in the load-bearing derivation. The framework is therefore self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Asymptotic analysis yields a closed nonlinear theory whose four regimes exhaust all possible combinations of the three heterogeneity parameters
Reference graph
Works this paper leans on
-
[1]
K. J. Cheung and S. Horne-Badovinac. Collective cell mi- gration modes in development, tissue repair and cancer. Nature Reviews Molecular Cell Biology, 26(10):741–758,
-
[2]
doi:10.1038/s41580-025-00858-9
-
[3]
Z. Wu and M. Wong. Collective cell migration across scales: A systems perspective.Seminars in Cell & Developmental Biology, 173:103628, 2025. doi:10.1016/j.semcdb.2025.103628
-
[4]
S. J. Stehbens, E. Scarpa, et al. Perspectives in collective cell migration – moving forward.Journal of Cell Science, 137(12):jcs261549, 2024. doi:10.1242/jcs.261549
-
[5]
J. Alanko, M. C. Uçar, et al. CCR7 acts as both a sensor and a sink for CCL19 to coordinate collective leukocyte migration.Science Immunology, 8(87):eadc9584, 2023. doi:10.1126/sciimmunol.adc9584
-
[6]
E. Donà, J. D. Barry, et al. Directional tissue migration through a self-generated chemokine gradient.Nature, 503(7475):285–289, 2013. doi:10.1038/nature12635
-
[7]
H. Z. Ford, G. L. Celora, et al. Pattern forma- tion along signaling gradients driven by active droplet behavior of cell swarms.Proceedings of the Na- tional Academy of Sciences, 122(21):e2419152122, 2025. doi:10.1073/pnas.2419152122
-
[8]
A. Kiran,P. A. Thomason, et al. Macrophage chemotaxis steered by complex self-generated gradients of comple- ment C5a.bioRxiv, 2025. doi:10.1101/2025.11.05.686842. 9
-
[9]
G. Malet-Engra, W. Yu, et al. Collective Cell Motil- ity Promotes Chemotactic Prowess and Resistance to Chemorepulsion.Current Biology, 25(2):242–250, 2015. doi:10.1016/j.cub.2014.11.030
-
[10]
L. Tweedy, P. A. Thomason, et al. Seeing around corners: Cells solve mazes and respond at a distance using at- tractant breakdown.Science, 369(6507):eaay9792, 2020. doi:10.1126/science.aay9792
-
[11]
M. C. Uçar, Z. Alsberga, et al. Self-generated chemotaxis of mixed cell populations.Proceed- ings of the National Academy of Sciences of the United States of America, 122(34):e2504064122, 2025. doi:10.1073/pnas.2504064122
-
[12]
L. Tweedy and R. H. Insall. Self-Generated Gradi- ents Yield Exceptionally Robust Steering Cues.Fron- tiers in Cell and Developmental Biology, 8:133, 2020. doi:10.3389/fcell.2020.00133
-
[13]
R. McLennan, L. Dyson, et al. Multiscale mecha- nisms of cell migration during development: theory and experiment.Development, 139(16):2935–2944, 2012. doi:10.1242/dev.081471
-
[14]
X. Fu, S. Kato, et al. Spatial self-organization re- solves conflicts between individuality and collective migration.Nature Communications, 9(1):2177, 2018. doi:10.1038/s41467-018-04539-4
-
[15]
E. F. Keller and L. A. Segel. Traveling bands of chemo- tactic bacteria: A theoretical analysis.Journal of Theo- retical Biology, 30(2):235–248, 1971. doi:10.1016/0022- 5193(71)90051-8
-
[16]
W. J. M. Ridgway, M. P. Dalwadi, et al. Motility- Induced Phase Separation Mediated by Bacterial Quo- rum Sensing.Physical Review Letters, 131(22):228302,
-
[17]
doi:10.1103/PhysRevLett.131.228302
-
[18]
M. M. Salek, F. Carrara, et al. Bacterial chemotaxis in a microfluidic T-maze reveals strong phenotypic hetero- geneity in chemotactic sensitivity.Nature Communica- tions, 10:1877, 2019. doi:10.1038/s41467-019-09521-2
-
[19]
H. H. Mattingly and T. Emonet. Collective behavior andnongeneticinheritanceallowbacterialpopulationsto adapt to changing environments.Proceedings of the Na- tional Academy of Sciences, 119(26):e2117377119, 2022. doi:10.1073/pnas.2117377119
-
[20]
S. Gude, E. Pinçe, et al. Bacterial coexistence driven by motility and spatial competition.Nature, 578(7796):588– 592, 2020. doi:10.1038/s41586-020-2033-2
-
[21]
Y. Bai, C. He, et al. Spatial modulation of individual behaviors enables an ordered structure of diverse pheno- types during bacterial group migration.eLife, 10:e67316,
-
[22]
doi:10.7554/eLife.67316
-
[23]
F. R. Macfarlane, T. Lorenzi, et al. The impact of phe- notypic heterogeneity on chemotactic self-organisation. Bulletin of Mathematical Biology, 84(12):143, 2022. doi:10.1007/s11538-022-01099-z
-
[24]
Phenotype structuring in collective cell migration: a tu- torial of mathematical models and methods
T. Lorenzi, K. J. Painter, et al. Phenotype structuring in collective cell migration: a tutorial of mathematical models and methods.Journal of Mathematical Biology, 90(6):61, 2025. doi:10.1007/s00285-025-02223-y
-
[25]
V. Freingruber, T. Lorenzi, et al. Trait-structured chemo- taxis: exploring ligand-receptor dynamics and travelling wave properties in a Keller–Segel model.Nonlinearity, 38(10):105006, 2025. doi:10.1088/1361-6544/ae03ff
-
[26]
Watts, C
M. Watts, C. Falco, et al. Patterns of collective migration in heterogeneous, self-generated chemotaxis. [Manuscript in preparation], 2026
2026
-
[27]
L. C. Evans.Partial differential equations, volume 19 of Graduate studies in mathematics. American Mathemati- cal Society, second edition, 2010
2010
-
[28]
M. J. Ablowitz and A. Zeppetella. Explicit solu- tions of Fisher’s equation for a special wave speed. Bulletin of Mathematical Biology, 41(6):835–840, 1979. doi:10.1007/BF02462380
-
[29]
C. M. Bender and S. A. Orszag.Advanced Mathematical Methods for Scientists and Engineers I. Springer, New York, NY, 1999
1999
-
[30]
J. Canosa. On a nonlinear diffusion equation describing population growth.IBM Journal of Research and Devel- opment, 17(4):307–313, 1973. doi:10.1147/rd.174.0307
-
[31]
D. D. Patel, W. Koopmann, et al. Chemokines have diverse abilities to form solid phase gra- dients.Clinical Immunology, 99(1):43–52, 2001. doi:10.1006/clim.2000.4997
-
[32]
T. V. Phan, H. H. Mattingly, et al. Direct measurement of dynamic attractant gradients reveals breakdown of the Patlak–Keller–Segel chemotaxis model.Proceedings of the National Academy of Sciences, 121(3):e2309251121,
-
[33]
doi:10.1073/pnas.2309251121
-
[34]
G. L. Celora, B. J. Walker, et al. Chemotaxis of cell ag- gregates: morphology and dynamics of migrating active droplets.arXiv, 2026. doi:10.48550/arXiv.2602.20088
-
[35]
T. J. Jewell, S. W. S. Johnson, et al. Cell-cell adhesion cannot sustain extended follower streams in a minimal non-local model of leader-follower migration.arXiv, 2026. doi:10.48550/arXiv.2604.15283
work page internal anchor Pith review Pith/arXiv arXiv doi:10.48550/arxiv.2604.15283 2026
-
[36]
E. Lawson-Keister and M. L. Manning. Collective chemotaxis in a Voronoi model for confluent clus- ters.Biophysical Journal, 121(23):4624–4634, 2022. doi:10.1016/j.bpj.2022.10.029. 10
-
[37]
L. Rossetti, S. Grosser, et al. Optogenetic generation of leader cells reveals a force–velocity relation for collective cell migration.Nature Physics, 20(10):1659–1669, 2024. doi:10.1038/s41567-024-02600-2
-
[38]
D. P. Panigrahi, G. L. Celora, et al. Intermit- tent attachments form three-dimensional cell aggre- gates with emergent fluid properties.bioRxiv, 2025. doi:10.1101/2025.09.24.678186
-
[39]
Dolak and C
Y. Dolak and C. Schmeiser. Kinetic models for chemo- taxis: Hydrodynamic limits and spatio-temporal mecha- nisms.Journal of Mathematical Biology, 51(6):595–615,
-
[40]
doi:10.1007/s00285-005-0334-6
-
[41]
J. Saragosti, V. Calvez, et al. Mathematical De- scription of Bacterial Traveling Pulses.PLOS Computational Biology, 6(8):e1000890, 2010. doi:10.1371/journal.pcbi.1000890
-
[42]
R. Alert. Cellular Sensing Governs the Stability of Chemotactic Fronts.Physical Review Letters, 128(14),
-
[43]
doi:10.1103/PhysRevLett.128.148101
-
[44]
T. Bhattacharjee, D. B. Amchin, et al. Chemotactic smoothing of collective migration.eLife, 11:e71226, 2022. doi:10.7554/eLife.71226
-
[45]
T. E. Fjelde, K. Xu, et al. Turing.jl: a general-purpose probabilistic programming language.ACM Transac- tions on Probabilistic Machine Learning, 1(3), 2025. doi:10.1145/3711897
-
[46]
M. D. Hoffman and A. Gelman. The No-U-Turn sam- pler: adaptively setting path lengths in Hamiltonian Monte Carlo.Journal of Machine Learning Research, 15(1):1593–1623, 2014. doi:10.5555/2627435.2638586
-
[47]
C. Rackauckas and Q. Nie. DifferentialEquations.jl–a performant and feature-rich ecosystem for solving dif- ferential equations in Julia.Journal of Open Research Software, 5(1), 2017. doi:10.5334/jors.151. 11 Supplementary Information Contents A Details of the SC model 1 A.1 Simulation of the dendritic/T-cell system . . . . . . . . . . . . . . . . . . . ....
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.