Kinetics of multistate DNA polymerases
Pith reviewed 2026-06-26 09:26 UTC · model grok-4.3
The pith
An iterative method computes the speed, error rate, and base-pair probabilities for T7 DNA polymerase replication more than a million times faster than simulations.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
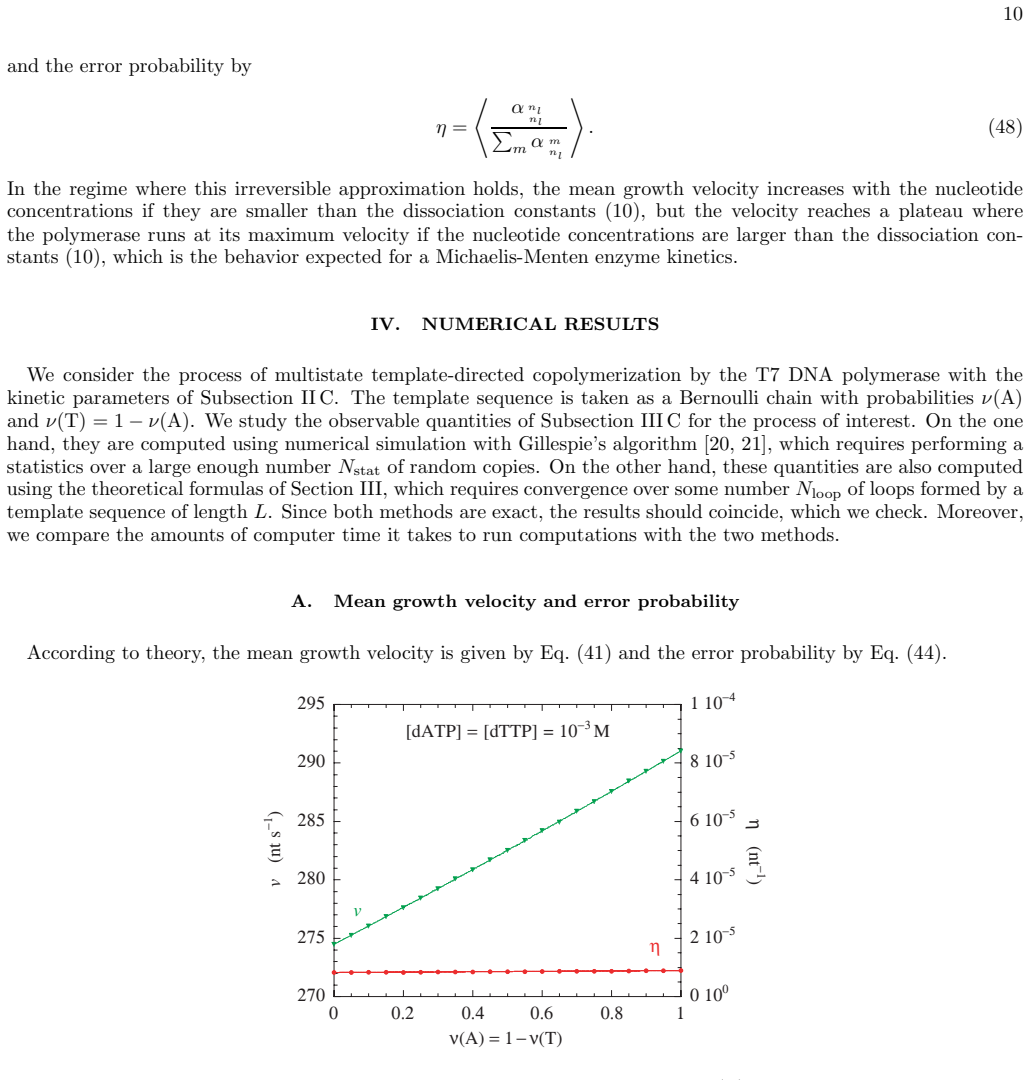
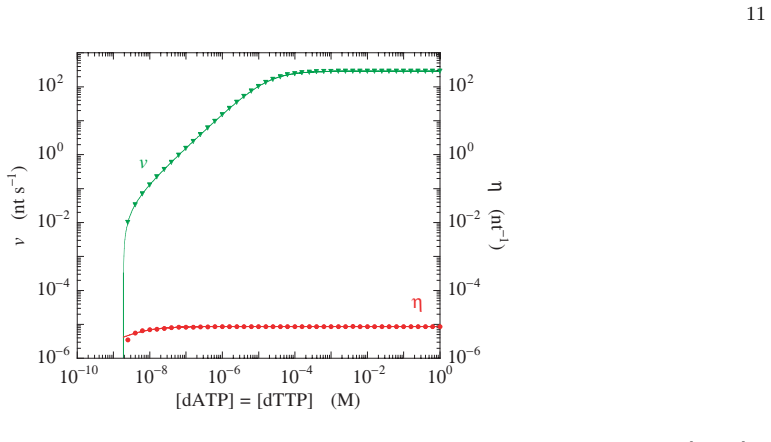
The iterative method previously developed for template-directed multistate copolymerization transfers to DNA replication by multistate polymerases. When applied to the two-state kinetic model for the T7 DNA polymerase, the method directly yields the mean velocity for growth of the copy along the template, the error probability of DNA replication, and the local probabilities of base-pair formation along the template sequence. The same calculations are more than a million times faster than usual numerical simulation methods, and additional results follow in the approximation of homogenization of template heterogeneities.
What carries the argument
The iterative mathematical method for the kinetics of template-directed multistate copolymerization, which computes the required quantities by successive iteration rather than full trajectory simulation.
If this is right
- The mean velocity, error probability, and local base-pair probabilities become directly available for any given template sequence under the two-state model.
- The same quantities can be obtained for homogenized templates that average over sequence heterogeneities.
- The computational cost drops by more than six orders of magnitude compared with direct stochastic simulation of the same process.
- The approach extends in principle to other polymerases that can be represented by a finite number of structural states.
Where Pith is reading between the lines
- The efficiency gain could make it feasible to scan entire genomes for predicted local error hotspots under the model.
- Adding more structural states to the polymerase description would remain tractable as long as the iteration converges.
- The same framework might connect replication kinetics to observed mutation rates in T7 phage populations.
- Direct experimental tests could compare the predicted velocity versus dNTP concentration against measured values.
Load-bearing premise
The two-state kinetic model is an adequate representation of the T7 DNA polymerase structural states and the iterative method transfers to this replication system without further adjustments.
What would settle it
A measured mean replication velocity or error probability for T7 DNA polymerase that differs from the value obtained by applying the iterative method to the two-state model.
Figures
read the original abstract
In the present paper, we apply the iterative mathematical method previously developed for the kinetics of template-directed multistate copolymerization to the kinetics of DNA replication by polymerases having multiple structural states. In particular, we study a two-state kinetic model for the T7 DNA polymerase. We obtain the mean velocity for the growth of the copy along the template, the error probability of DNA replication by the polymerase, and the local probabilities of base-pair formation along the template sequence. Furthermore, we show that the iterative method is more than a million times faster than usual numerical simulation methods. Results are also obtained in the approximation of homogenization of template heterogeneities.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript applies a previously developed iterative method for the kinetics of template-directed multistate copolymerization to a two-state kinetic model of T7 DNA polymerase. It reports computation of the mean velocity of copy-strand growth along the template, the replication error probability, and local base-pair formation probabilities; it further claims the iterative method is more than 10^6 times faster than direct numerical simulation of the master equation and presents results under a homogenization approximation for template heterogeneities.
Significance. If the direct transfer of the iterative method holds without unstated boundary or processivity adjustments, the work supplies an efficient, exact route to steady-state kinetic observables for multistate polymerase models. The reported computational speedup constitutes a concrete practical advantage for exploring sequence-dependent replication fidelity, and the homogenization results provide a controlled approximation for heterogeneous templates.
major comments (2)
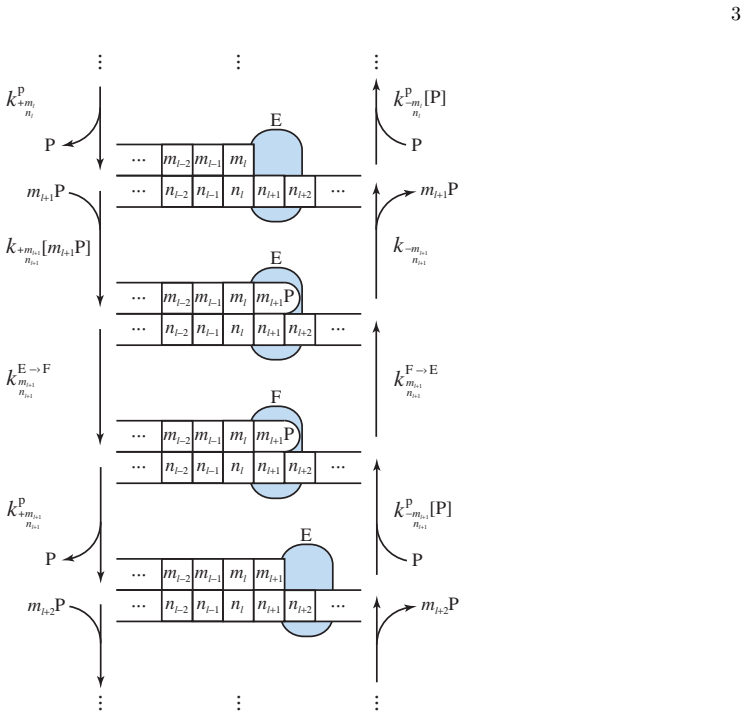
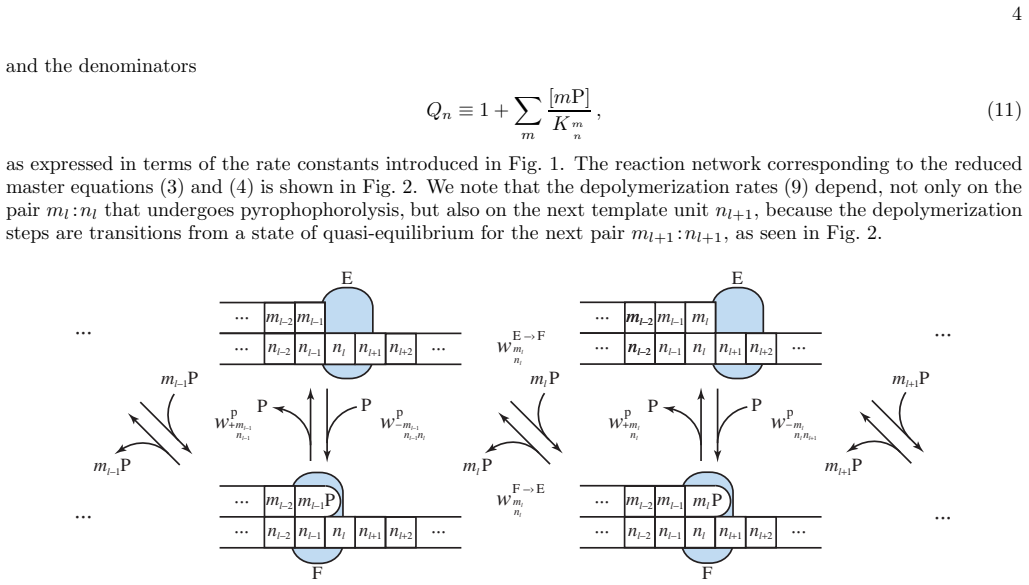
- [§2, §4] §2 (model definition) and §4 (application of iteration): the manuscript must explicitly verify that the two-state polymerase master equation (with sequence-dependent insertion and exonuclease rates) maps onto the copolymerization framework without additional termination or processivity boundary conditions; otherwise the claimed exactness of the mean velocity and error probability cannot be confirmed from the prior method alone.
- [§5] §5 (numerical results): the >10^6 speedup is presented as an empirical demonstration, but the manuscript should report the system size, number of Monte Carlo trajectories, and convergence criterion used for the direct simulation baseline so that the timing comparison is reproducible and not dependent on implementation details.
minor comments (3)
- [Abstract, §1] The abstract and introduction should cite the specific prior reference for the iterative method (including equation numbers) rather than describing it only as 'previously developed'.
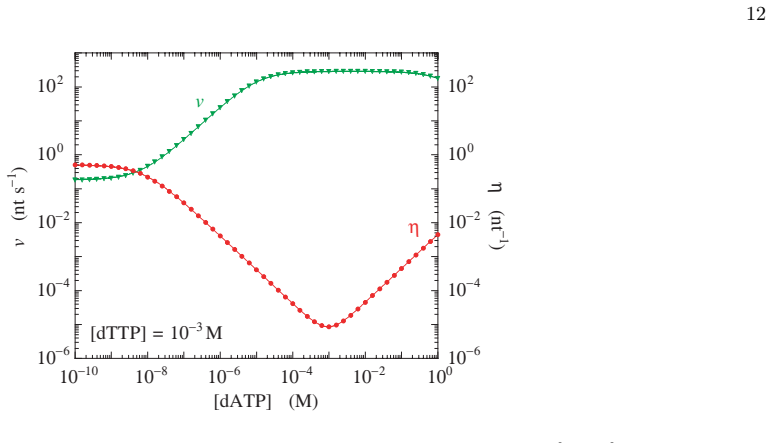
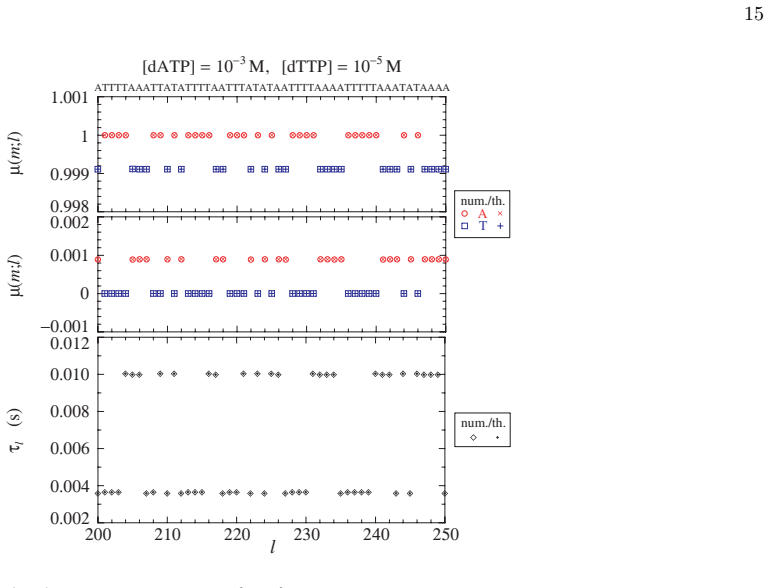
- [Figures 2-4] Figure captions for the velocity and error-probability plots should state the template length, the specific rate constants employed, and whether the plotted quantities are steady-state values or time averages.
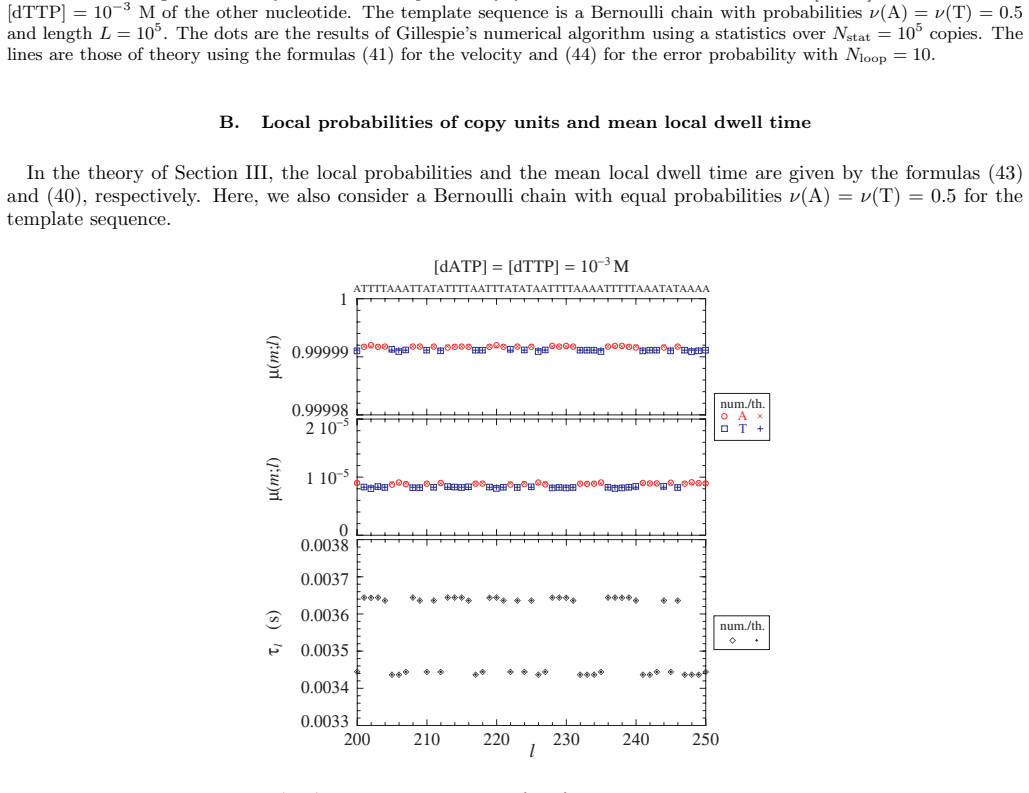
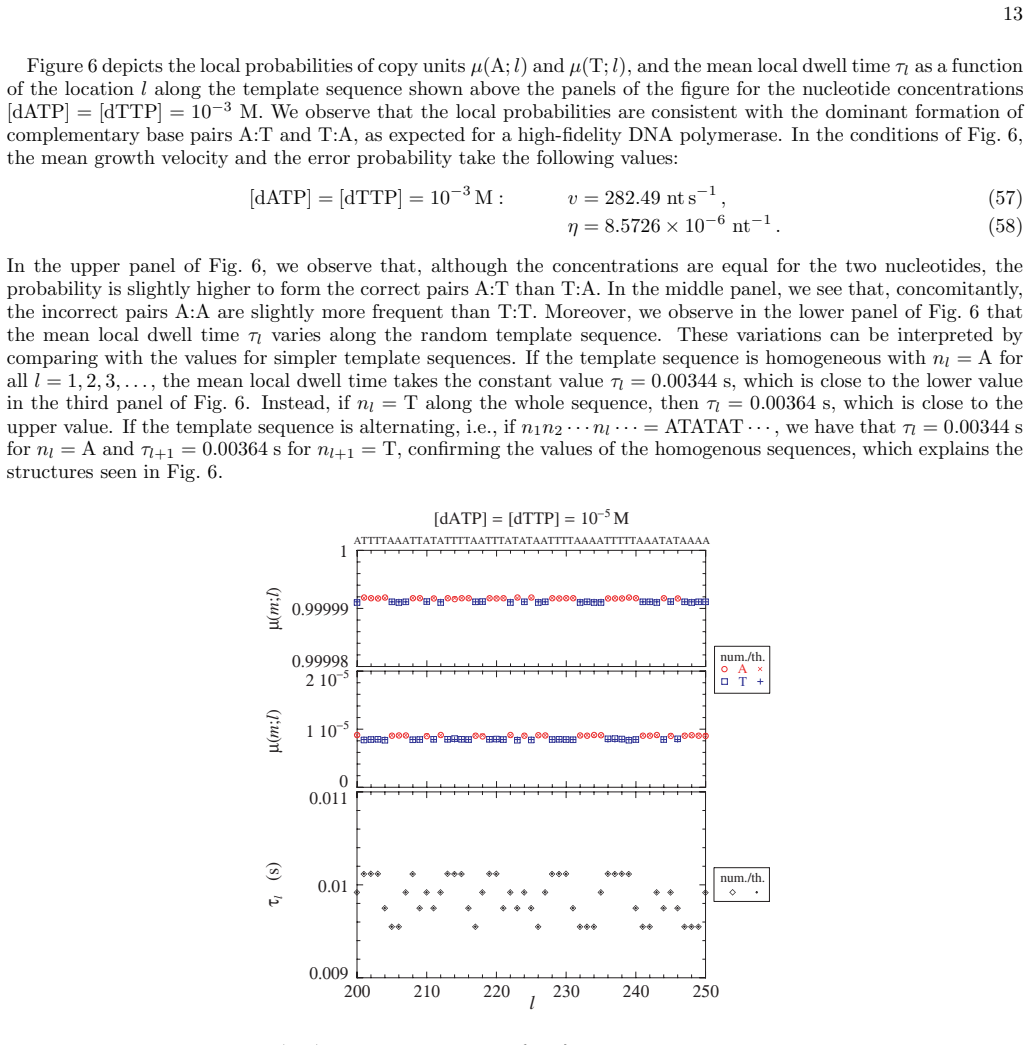
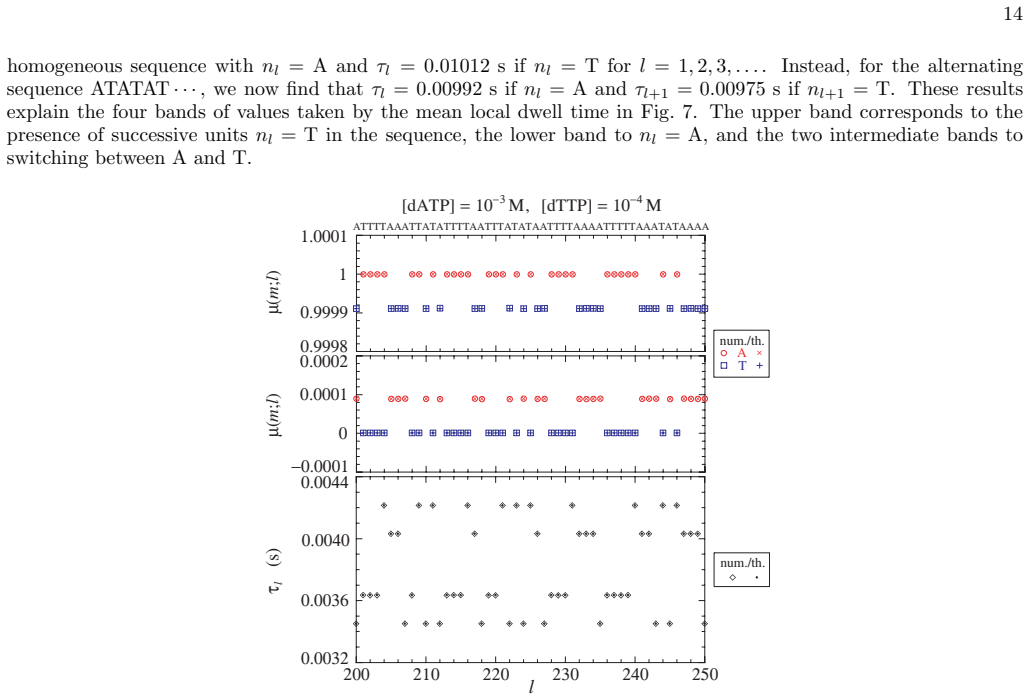
- [§3] Notation for the local base-pair probabilities p_i should be defined once and used consistently; the relation between these probabilities and the error probability should be stated explicitly.
Simulated Author's Rebuttal
We thank the referee for their careful review and constructive comments, which will improve the clarity and reproducibility of the manuscript. We address each major comment below and will revise accordingly.
read point-by-point responses
-
Referee: [§2, §4] §2 (model definition) and §4 (application of iteration): the manuscript must explicitly verify that the two-state polymerase master equation (with sequence-dependent insertion and exonuclease rates) maps onto the copolymerization framework without additional termination or processivity boundary conditions; otherwise the claimed exactness of the mean velocity and error probability cannot be confirmed from the prior method alone.
Authors: We agree that an explicit verification strengthens the claims. In the revised manuscript we will insert a dedicated paragraph (or short subsection) in §2 that derives the correspondence between the two-state T7 polymerase master equation (including the sequence-dependent insertion and exonuclease rates) and the multistate copolymerization framework of the prior work. We will confirm that the boundary conditions remain exactly those of the original iterative method, with no additional termination or processivity terms required. This addition will make the exactness of the velocity and error-probability results directly verifiable from the mapping alone. revision: yes
-
Referee: [§5] §5 (numerical results): the >10^6 speedup is presented as an empirical demonstration, but the manuscript should report the system size, number of Monte Carlo trajectories, and convergence criterion used for the direct simulation baseline so that the timing comparison is reproducible and not dependent on implementation details.
Authors: We accept this request for full reproducibility. The revised §5 will report (i) the template length used as system size, (ii) the exact number of Monte Carlo trajectories executed for the direct master-equation simulation, and (iii) the convergence criterion (e.g., relative change in velocity or error probability below a stated threshold) employed to terminate the runs. These parameters will be stated explicitly so that the timing comparison can be reproduced independently of implementation details. revision: yes
Circularity Check
No significant circularity
full rationale
The paper applies an iterative method from prior work to a two-state T7 polymerase model to compute mean velocity, error probability, and local base-pair probabilities, while demonstrating computational speedup versus direct simulation. The derivation chain consists of model definition followed by application of the established iteration; no equations or results within this manuscript reduce by construction to fitted parameters, self-referential definitions, or unverified self-citations. The prior method is treated as an external tool whose validity is independent of the present application, and all reported quantities are obtained within the explicit model without renormalization or ansatz smuggling.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption The iterative mathematical method previously developed for template-directed multistate copolymerization applies without modification to multistate DNA polymerases.
Reference graph
Works this paper leans on
-
[1]
In this stationary regime, the mean length of the copy remains finite up to nucleotide concentratio n values corresponding to the onset of growth, as determined in Appendix C
Stationary regime If the nucleotide concentrations are too small, the growth of the c opy is stalled. In this stationary regime, the mean length of the copy remains finite up to nucleotide concentratio n values corresponding to the onset of growth, as determined in Appendix C. For larger values of the nucleotide conc entrations, the growth of the copy beco...
-
[2]
Regime of sublinear growth in time As explained in Appendix C, if the template sequence is disordered, th ere is a regime where the mean length has a sublinear growth in time as ⟨l⟩t ∼ tγ with an exponent 0 <γ < 1, where γ = 0 at the onset of growth and γ = 1 at the threshold of the steady-growth regime
-
[3]
Steady-growth regime In this regime, the mean length of the copy grows linearly in time like ⟨l⟩t ≃ vt with a positive mean growth velocityv >0 given by Eq. (41). Close to the threshold γ = 1, the depolymerization rates wp − mlnlnl+1 continue to play a significant role in determining the velocity v. However, their role becomes negligible away beyond the th...
-
[4]
Generalities Here, we determine the kinetic parameters of Table I for the two-s tate model of T7 DNA polymerase using the experimental data of Refs. [4–6]. These data are measured with kin etic methods for a given template sequence, a given initial sequence for the copy, and a given nucleotide concentr ation [dNTP], which is either [dATP] or [dTTP]. These...
-
[5]
4 of Ref
The parameter values for dATP:T According to the data in Table 2 on p. 4 of Ref. [6], the constants (B2 ) and (B3) take the following values, kcat = 280 s − 1 and Km = 1. 8 × 10− 5 M (B5) for this correct pair. Furthermore, the values k2 = 6500 s− 1 and k− 2 = 1. 7 s− 1 (B6) are reported on p. 12 of Ref. [6]. Therefore, inverting Eq. (B2), w e obtain k3 =...
-
[6]
8 × 10− 5 M (B9) according to Table 2 on p
The parameter values for dTTP:A For this other correct pair, we have that kcat = 297 s − 1 and Km = 1. 8 × 10− 5 M (B9) according to Table 2 on p. 4 of Ref. [6]. Moreover, we assume the sam e values k2 = 6500 s− 1 and k− 2 = 1. 7 s− 1 (B10) for the two correct pairs. In a similar way as in Eqs. (B7) and (B8), w e here find that k3 = 311 s− 1 and K1 = 3. 92...
-
[7]
14 of Ref
The parameter values for dTTP:T Table 6 on p. 14 of Ref. [6] gives the following parameter values in the c ase of two-step nucleotide binding: K1 = 9. 3 × 10− 3 M, k 2 = 170 s − 1, and k− 2 = 340 s− 1. (B12) Now, taking the value k3 = 2. 4 s− 1 and using Eq. (B4), we find that kcat/K m = 128 M − 1 s− 1, which agrees with the experimental value kcat/K m = 1...
-
[8]
4 of Ref
The parameter values for dATP:A The lower bound on the value of kcat given in Table 2 on p. 4 of Ref. [6] for the pair dATP:A is about 2 . 6 times larger than for dTTP:T, which suggests that the rate constant k3 is here larger in the same proportion. Accordingly, we here suppose that k3 = 6. 2 s− 1. In addition, we assume that the rate constants of the re...
-
[9]
Pyrophosphorolysis The experimental value (13) of the pyrophosphorolysis constant is given in Fig. 4 on p. 7 of Ref. [5] for the correct pair dATP:T. This value implies that the nucleotide concentration at ze ro velocity should be given by [dNTP]v=0 ≃ K1k− 2 k2KP [P] = 1. 9 × 10− 9 M (B15) for [P] = 10 − 4 M, which is consistent with Eq. (78) and with pre...
-
[10]
Gaspard, Kinetics of template-directed multistate copolymerizati on, companion paper, preprint (2026)
P. Gaspard, Kinetics of template-directed multistate copolymerizati on, companion paper, preprint (2026)
2026
-
[11]
Tsai and K
Y.-C. Tsai and K. A. Johnson, A new paradigm for DNA polymerase specificity , Biochem. 45, 9675–9687 (2006)
2006
-
[12]
K. A. Johnson, The kinetic and chemical mechanism of high-fidelity DNA poly merases, Biochim. Biophys. Acta 1804, 1041–1048 (2010)
2010
-
[13]
T. L. Dangerfield and K. A. Johnson, Optimized incorporation of an unnatural fluorescent amino a cid affords measurement of conformational dynamics governing high-fidelity DNA rep lication, J. Biol. Chem. 295, 17265–17820 (2020)
2020
-
[14]
T. L. Dangerfield and K. A. Johnson, Conformational dynamics during high-fidelity DNA replicat ion and translocation defined using a DNA polymerase with a fluorescent artificial am ino acid , J. Biol. Chem. 296, 100143 (2021)
2021
-
[15]
T. L. Dangerfield, S. Kirmizialtin, and K. A. Johnson, Conformational dynamics during misincorporation and mism atch extension defined using a DNA polymerase with a fluorescent ar tificial amino acid , J. Biol. Chem. 298, 101451 (2022)
2022
-
[16]
Michaelis and M
L. Michaelis and M. L. Menten, Die Kinetik der Invertinwirkung , Biochem. Z. 49, 333–369 (1913); English translation in Ref. [8]
1913
-
[17]
K. A. Johnson and R. S. Goody, The Original Michaelis Constant: Translation of the 1913 Mi chaelis–Menten Paper , Biochem. 50, 8264–8269 (2011)
1913
-
[18]
Stryer, Biochemistry, 2nd Edition (Freeman and Co., San Francisco, 1981)
L. Stryer, Biochemistry, 2nd Edition (Freeman and Co., San Francisco, 1981)
1981
-
[19]
A. L. Lehninger, Principles of Biochemistry (Worth Publishers, New York, 1982)
1982
-
[20]
S. S. Patel, I. Wong, and K. A. Johnson, Pre-Steady-State Kinetic Analysis of Processive DNA Repli cation Including Complete Characterization of an Exonuclease-Deficient Mut ant, Biochem. 30, 511–525 (1991)
1991
-
[21]
I. Wong, S. S. Patel, and K. A. Johnson, An Induced-Fit Kinetic Mechanism for DNA Replication Fidel ity: Direct Mea- surement by Single-Turnover Kinetics , Biochem. 30, 526–537 (1991)
1991
-
[22]
K. A. Johnson, Conformational Coupling in DNA Polymerase Fidelity , Annu. Rev. Biochem. 62, 685–713 (1993)
1993
-
[23]
J. Ahn, B. G. Werneburg, and M.-D. Tsai, DNA Polymerase β : Structure–Fidelity Relationship from Pre-Steady-State Kinetic Analyses of All Possible Correct and Incorrect Base Pairs for Wild Type and R283A Mutant , Biochem. 36, 1100– 1107 (1997)
1997
-
[24]
K. A. Fiala and Z. Suo, Pre-Steady-State Kinetic Studies of the Fidelity of Sulfol obus solfataricus P2 DNA Polymerase IV , Biochem. 43, 2106–2115 (2004)
2004
-
[25]
H. R. Lee and K. A. Johnson, Fidelity of the Human Mitochondrial DNA Polymerase , J. Biol. Chem. 281, 36236–36240 (2006)
2006
-
[26]
Zhang, J
L. Zhang, J. A. Brown, S. A. Newmister, and Z. Suo, Polymerization Fidelity of a Replicative DNA Polymerase fr om the Hyperthermophilic Archaeon Sulfolobus solfataricus P2 , Biochem. 48, 7492–7501 (2009)
2009
-
[27]
R. J. Bauer, M. T. Begley, and M. A. Trakselis, Kinetics and Fidelity of Polymerization by DNA Polymerase I II from Sulfolobus solfataricus , Biochem. 51, 1996–2007 (2012)
1996
-
[28]
K. M. Schermerhorn and A. F. Gardner, Pre-steady-state Kinetic Analysis of a Family D DNA Polymer ase from Thermo- coccus sp. 9◦N Reveals Mechanisms for Archaeal Genomic Replication and M aintenance, J. Biol. Chem. 290, 21800–21810 (2015)
2015
-
[29]
D. T. Gillespie, A general method for numerically simulating the stochastic time evolution of coupled chemical reactions , J. Comput. Phys. 22, 403–434 (1976)
1976
-
[30]
D. T. Gillespie, Exact stochastic simulation of coupled chemical reactions , J. Phys. Chem. 81, 2340–2361 (1977)
1977
-
[31]
M. F. Barnsley and S. Demko, Iterated function systems and the global construction of fr actals, Proc. R. Soc. Lond. A 399, 243–275 (1985)
1985
-
[32]
Gaspard, Template-directed copolymerization, random walks along d isordered tracks, and fractals , Phys
P. Gaspard, Template-directed copolymerization, random walks along d isordered tracks, and fractals , Phys. Rev. Lett. 117, 238101 (2016). 23
2016
-
[33]
Gaspard, Iterated function systems for DNA replication , Phys
P. Gaspard, Iterated function systems for DNA replication , Phys. Rev. E 96, 042403 (2017)
2017
-
[34]
J. K. Heinonen, Biological Role of Inorganic Pyrophosphate (Springer, New York, 2001)
2001
-
[35]
K. A. Johnson, Transient-state kinetic analysis of enzyme reaction pathw ays, The Enzymes XX, 1–61 (1992)
1992
-
[36]
Gaspard, Kinetics and thermodynamics of exonuclease-deficient DNA p olymerases, Phys
P. Gaspard, Kinetics and thermodynamics of exonuclease-deficient DNA p olymerases, Phys. Rev. E 93, 042419 (2016)
2016
-
[37]
Andrieux and P
D. Andrieux and P. Gaspard, Molecular information processing in nonequilibrium copol ymerizations, J. Chem. Phys. 130, 014901 (2009)
2009
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.