Feature leakage and the identifiability of direct-dependency entropy models of neural activity
Pith reviewed 2026-06-28 12:05 UTC · model grok-4.3
The pith
Direct entropy models of neural activity measure prediction under sampled inputs rather than identifying response mechanisms.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
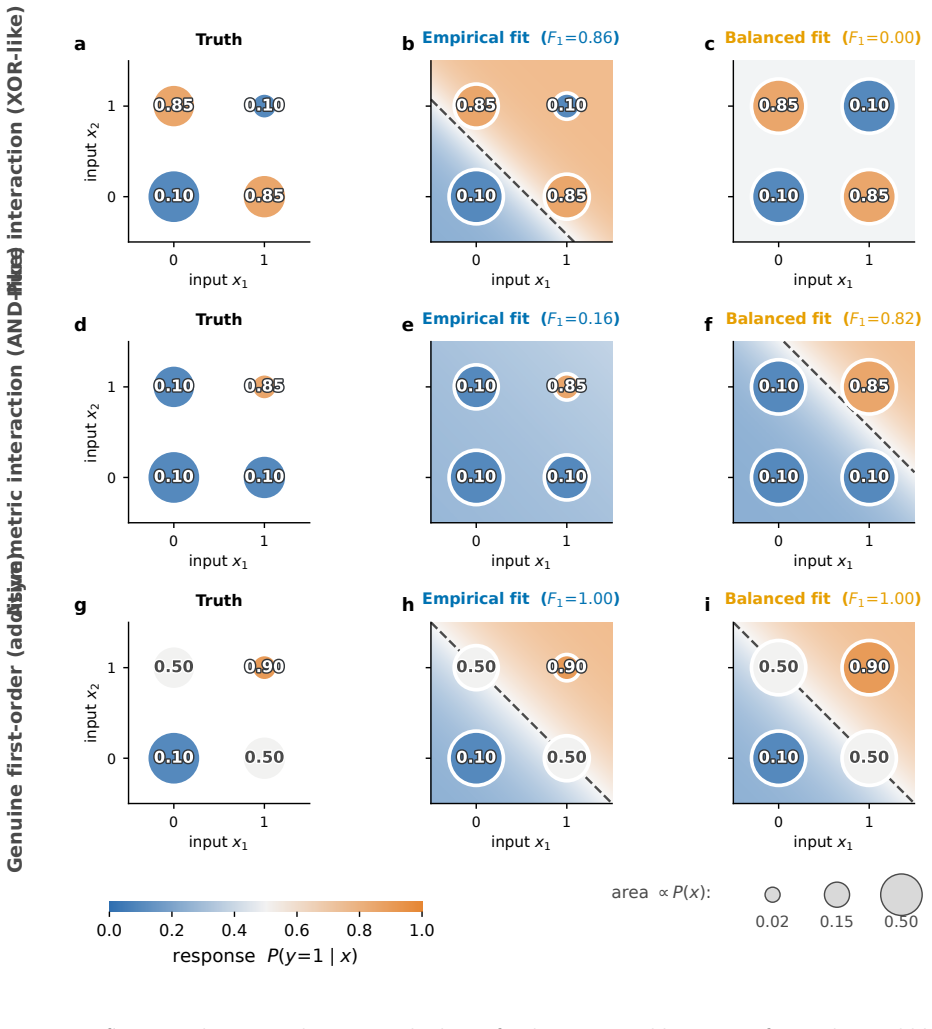
For conditional maximum-entropy models that match output rates and pairwise output-input coactivities, the entropy explained by a direct model is a prediction measure under the sampled input distribution, not a mechanism-identification test. A restricted MaxEnt fit is an information projection: omitted interaction, temporal, or hidden-state terms can be absorbed into fitted first-order parameters whenever they are correlated with the included sufficient statistics. For sparse correlated binary inputs, this absorption has an explicit coskewness form. Diagnostics that hold P(y|x) fixed while changing P(x) correctly classify higher-order responses that masquerade as direct under empirical sampl
What carries the argument
State reweighting that holds the conditional response P(y|x) fixed while varying the input distribution P(x) to expose absorption of omitted terms into first-order parameters.
If this is right
- Apparent first-order fits can arise from higher-order responses when input sampling induces absorption into the fitted parameters.
- State reweighting isolates distribution-dependent fits from those that recover the underlying response rule.
- In CA1 data, many tables stable under empirical weights become sensitive once the input distribution is balanced.
- Raw coactivity predictions and entropy-explained fractions should be treated as in-distribution performance metrics only.
Where Pith is reading between the lines
- The same absorption mechanism may affect other projection-based models of neural population activity whenever inputs are correlated.
- Controlled reweighting experiments could be used to test whether apparent simplicity in other brain areas reflects computation or sampling.
- Quantifying the coskewness term explicitly might allow bias correction in existing first-order fits without new recordings.
Load-bearing premise
That correlations between omitted higher-order terms and the included first-order statistics remain negligible under the observed input distribution.
What would settle it
A ground-truth simulation with a known higher-order response rule that continues to receive first-order classifications after state reweighting is applied.
Figures




read the original abstract
Biological neurons receive thousands of synaptic inputs on branching, electrically excitable dendrites, yet population activity is often modeled with direct input-output rules in which each input contributes independently to a scalar drive. We study what successful prediction by such models does, and does not, reveal about neural computation. For conditional maximum-entropy models that match output rates and pairwise output-input coactivities, the entropy explained by a direct model is a prediction measure under the sampled input distribution, not a mechanism-identification test. A restricted MaxEnt fit is an information projection: omitted interaction, temporal, or hidden-state terms can be absorbed into fitted first-order parameters whenever they are correlated with the included sufficient statistics. For sparse correlated binary inputs, this absorption has an explicit coskewness form. We introduce diagnostics that separate in-distribution prediction from recovery of the response rule: state reweighting that holds P(y|x) fixed while changing P(x), conditional log-odds contrasts for local additivity, and temporal leakage controls. In ground-truth simulations, purely higher-order responses can pass first-order entropy and raw coactivity tests under leakage-prone sampling, but are correctly classified after reweighting. Applied to selected, leakage-enriched local tables from CA1 hippocampal recordings, approximately half of tables that appear first-order under empirical weights become distribution-sensitive under balanced reweighting, far above a matched additive-surrogate null. Thus direct entropy-explained fractions and raw coactivity predictions should be interpreted as predictions under the observed state distribution, not as evidence that mechanisms outside the direct model are absent or small.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that for conditional maximum-entropy models matching output rates and pairwise output-input coactivities, the entropy explained by a direct model is an in-distribution prediction measure under the sampled input distribution rather than a mechanism-identification test. Omitted higher-order, temporal, or hidden-state terms can be absorbed into fitted first-order parameters; for sparse correlated binary inputs this absorption takes an explicit coskewness form. The authors introduce state reweighting (holding P(y|x) fixed while altering P(x)), conditional log-odds contrasts, and temporal leakage controls as diagnostics. Ground-truth simulations show that purely higher-order responses can pass first-order tests under leakage-prone sampling but are correctly reclassified after reweighting. Applied to selected leakage-enriched local tables from CA1 recordings, approximately half of tables that appear first-order under empirical weights become distribution-sensitive under balanced reweighting, exceeding a matched additive-surrogate null.
Significance. If the central mathematical and diagnostic claims hold, the work supplies a precise caution for the interpretation of direct-dependency MaxEnt models in systems neuroscience. The explicit linkage of entropy explained to information projection, the coskewness absorption term, and the reweighting procedure that isolates response-rule recovery from sampling effects constitute a useful methodological advance. The ground-truth simulations and the quantitative CA1 finding provide concrete evidence that raw coactivity predictions should not be read as evidence against omitted mechanisms.
major comments (2)
- [Results, CA1 application] CA1 application paragraph: the claim that 'approximately half of tables' become distribution-sensitive is presented without the total number of tables, exact fraction, confidence intervals, or a statistical comparison (e.g., p-value) to the additive-surrogate null. This quantitative result is load-bearing for the empirical support of the central claim.
- [Theory / Methods on coskewness] Theory section on absorption: although the abstract asserts an explicit coskewness form for the leakage term, the derivation or the explicit equation is not supplied in the visible text; without it the absorption statement cannot be verified independently and remains a load-bearing step for the identifiability argument.
minor comments (1)
- [Methods, state reweighting] Notation for the reweighting procedure: the distinction between the empirical input distribution and the reweighted distribution could be clarified with a small table or explicit formulas for the weights.
Simulated Author's Rebuttal
We thank the referee for the detailed and constructive report. The two major comments identify genuine gaps in quantitative reporting and derivation visibility that we will address directly in revision. We respond to each point below.
read point-by-point responses
-
Referee: [Results, CA1 application] CA1 application paragraph: the claim that 'approximately half of tables' become distribution-sensitive is presented without the total number of tables, exact fraction, confidence intervals, or a statistical comparison (e.g., p-value) to the additive-surrogate null. This quantitative result is load-bearing for the empirical support of the central claim.
Authors: We agree that the current text omits these essential details. In the revised manuscript we will state the total number of local tables analyzed, the exact fraction (and count) that become distribution-sensitive under reweighting, bootstrap or analytic confidence intervals on that fraction, and a direct statistical comparison (p-value) against the matched additive-surrogate null. These numbers will be added to the CA1 results paragraph and, if space permits, to a supplementary table. revision: yes
-
Referee: [Theory / Methods on coskewness] Theory section on absorption: although the abstract asserts an explicit coskewness form for the leakage term, the derivation or the explicit equation is not supplied in the visible text; without it the absorption statement cannot be verified independently and remains a load-bearing step for the identifiability argument.
Authors: We acknowledge that the explicit equation and its derivation are not shown in the main text. In the revised version we will insert, in the Theory section immediately after the statement that absorption takes a coskewness form, the full derivation together with the resulting closed-form expression for the first-order parameter shift induced by omitted third-order input statistics. The derivation will be self-contained and will reference only the sparse-binary-input assumptions already stated in the manuscript. revision: yes
Circularity Check
No significant circularity identified
full rationale
The paper's central claim follows from the standard definition of conditional maximum-entropy models as information projections onto moment-matching distributions, but this is invoked only as clarification to motivate independent diagnostics (state reweighting that fixes P(y|x) while varying P(x), conditional log-odds contrasts, and temporal controls). Ground-truth simulations and the CA1 application supply external validation that is not reducible to the definitional property. No load-bearing step equates a derived quantity to its inputs by construction, and no self-citation chain is load-bearing.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption A restricted MaxEnt fit is an information projection onto the included sufficient statistics
Reference graph
Works this paper leans on
-
[1]
Simple input-output dependencies explain neuronal activity,
C. W. Lynn, “Simple input-output dependencies explain neuronal activity,”Nature Physics, https://doi.org/10.1038/s41567-026-03306-3 (2026). 20
-
[2]
Information theory and statistical mechanics,
E. T. Jaynes, “Information theory and statistical mechanics,”Physical Review106, 620–630 (1957)
1957
-
[3]
I-divergence geometry of probability distributions and minimization problems,
I. Csisz´ ar, “I-divergence geometry of probability distributions and minimization problems,” Annals of Probability3, 146–158 (1975)
1975
-
[4]
Amari,Information Geometry and Its Applications(Springer, 2016)
S.-I. Amari,Information Geometry and Its Applications(Springer, 2016)
2016
-
[5]
Graphical models, exponential families, and variational inference,
M. J. Wainwright and M. I. Jordan, “Graphical models, exponential families, and variational inference,”Foundations and Trends in Machine Learning1, 1–305 (2008)
2008
-
[6]
The behavior of maximum likelihood estimates under nonstandard conditions,
P. J. Huber, “The behavior of maximum likelihood estimates under nonstandard conditions,” inProceedings of the Fifth Berkeley Symposium on Mathematical Statistics and Probability, Vol. 1, 221–233 (1967)
1967
-
[7]
Maximum likelihood estimation of misspecified models,
H. White, “Maximum likelihood estimation of misspecified models,”Econometrica50, 1–25 (1982)
1982
-
[8]
Weak pairwise correlations imply strongly correlated network states in a neural population,
E. Schneidman, M. J. Berry II, R. Segev and W. Bialek, “Weak pairwise correlations imply strongly correlated network states in a neural population,”Nature440, 1007–1012 (2006)
2006
-
[9]
Searching for collective behavior in a large network of sensory neurons,
G. Tkaˇ cik, O. Marre, D. Amodei, E. Schneidman, W. Bialek and M. J. Berry II, “Searching for collective behavior in a large network of sensory neurons,”PLoS Computational Biology10, e1003408 (2014)
2014
-
[10]
Sparse low-order interaction network underlies a highly correlated and learnable neural population code,
E. Ganmor, R. Segev and E. Schneidman, “Sparse low-order interaction network underlies a highly correlated and learnable neural population code,”Proceedings of the National Academy of Sciences108, 9679–9684 (2011)
2011
-
[11]
Common input explains higher-order correlations and entropy in a simple model of neural population activity,
J. H. Macke, M. Opper and M. Bethge, “Common input explains higher-order correlations and entropy in a simple model of neural population activity,”Physical Review Letters106, 208102 (2011)
2011
-
[12]
Spatio-temporal correlations and visual signalling in a complete neuronal population,
J. W. Pillow et al., “Spatio-temporal correlations and visual signalling in a complete neuronal population,”Nature454, 995–999 (2008)
2008
-
[13]
Systematic errors in connectivity inferred from activity in strongly recurrent networks,
A. Das and I. R. Fiete, “Systematic errors in connectivity inferred from activity in strongly recurrent networks,”Nature Neuroscience23, 1286–1296 (2020)
2020
-
[14]
Minimal models of multidimensional computations,
J. D. Fitzgerald, L. C. Sincich and T. O. Sharpee, “Minimal models of multidimensional computations,”PLoS Computational Biology7, e1001111 (2011)
2011
-
[15]
Pairwise maximum entropy models for studying large biological systems: when they can work and when they can’t,
Y. Roudi, S. Nirenberg and P. E. Latham, “Pairwise maximum entropy models for studying large biological systems: when they can work and when they can’t,”PLoS Computational Biology5, e1000380 (2009)
2009
-
[16]
The impact of high-order interactions on the rate of synchronous discharge and information transmission in somatosensory cortex,
F. Montani, R. A. A. Ince, R. Senatore, E. Arabzadeh, M. E. Diamond and S. Panzeri, “The impact of high-order interactions on the rate of synchronous discharge and information transmission in somatosensory cortex,”Philosophical Transactions of the Royal Society A367, 3297–3310 (2009)
2009
-
[17]
Partitioning neuronal variability,
R. L. T. Goris, J. A. Movshon and E. P. Simoncelli, “Partitioning neuronal variability,”Nature Neuroscience17, 858–865 (2014). 21
2014
-
[18]
Latent dynamical variables produce signatures of spatiotemporal criticality in large biological systems,
M. C. Morrell, A. J. Sederberg and I. Nemenman, “Latent dynamical variables produce signatures of spatiotemporal criticality in large biological systems,”Physical Review Letters 126, 118302 (2021)
2021
-
[19]
Pyramidal neuron as two-layer neural network,
P. Poirazi, T. Brannon and B. W. Mel, “Pyramidal neuron as two-layer neural network,”Neuron 37, 989–999 (2003)
2003
-
[20]
Computational subunits in thin dendrites of pyramidal cells,
A. Polsky, B. W. Mel and J. Schiller, “Computational subunits in thin dendrites of pyramidal cells,”Nature Neuroscience7, 621–627 (2004)
2004
-
[21]
Dendritic computation,
M. London and M. H¨ ausser, “Dendritic computation,”Annual Review of Neuroscience28, 503–532 (2005)
2005
-
[22]
Dendritic spikes enhance stimulus selectivity in cortical neurons in vivo,
S. L. Smith, I. T. Smith, T. Branco and M. H¨ ausser, “Dendritic spikes enhance stimulus selectivity in cortical neurons in vivo,”Nature503, 115–120 (2013). 22 Supplementary Information A Theory and identifiability results a first-order family absorbed residual true response first-order fit Information projection 000 100 010 111 110 input state (x1x2x3) 0...
2013
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.