Separating wiring-specific from statistical control of dynamics in a complete connectome
Pith reviewed 2026-06-26 21:59 UTC · model grok-4.3
The pith
Coarse wiring statistics set the dynamical regime of the larval Drosophila connectome while precise connections control where activity flows and which circuits dominate.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
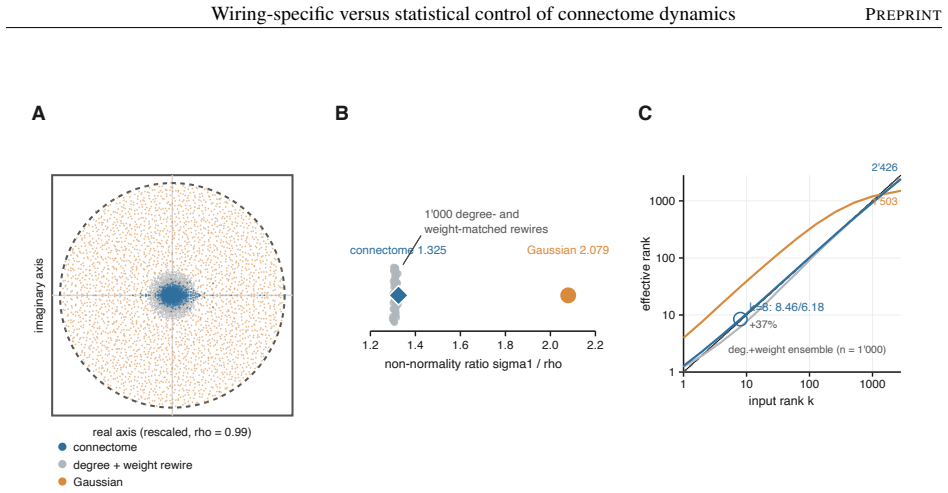
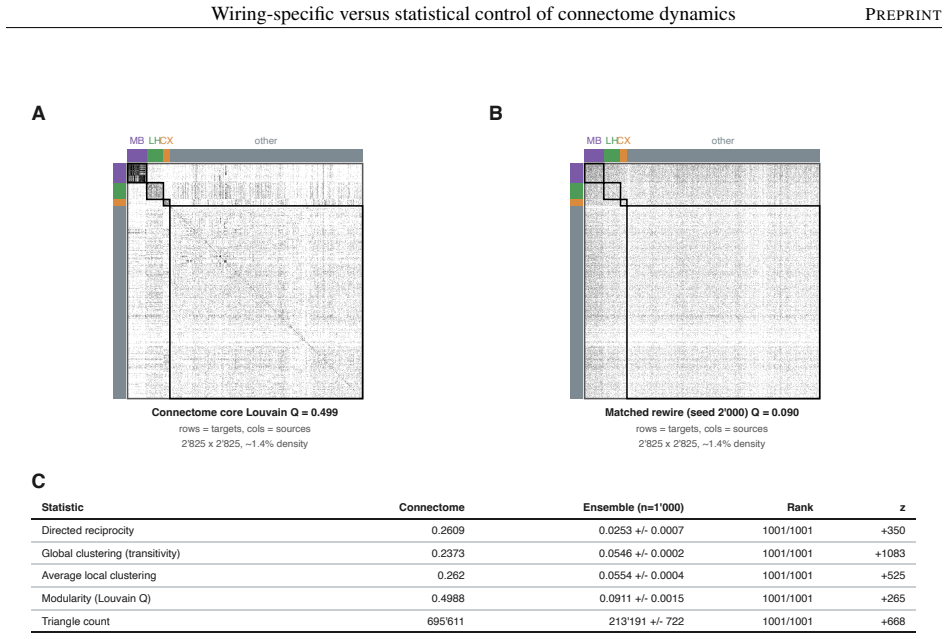
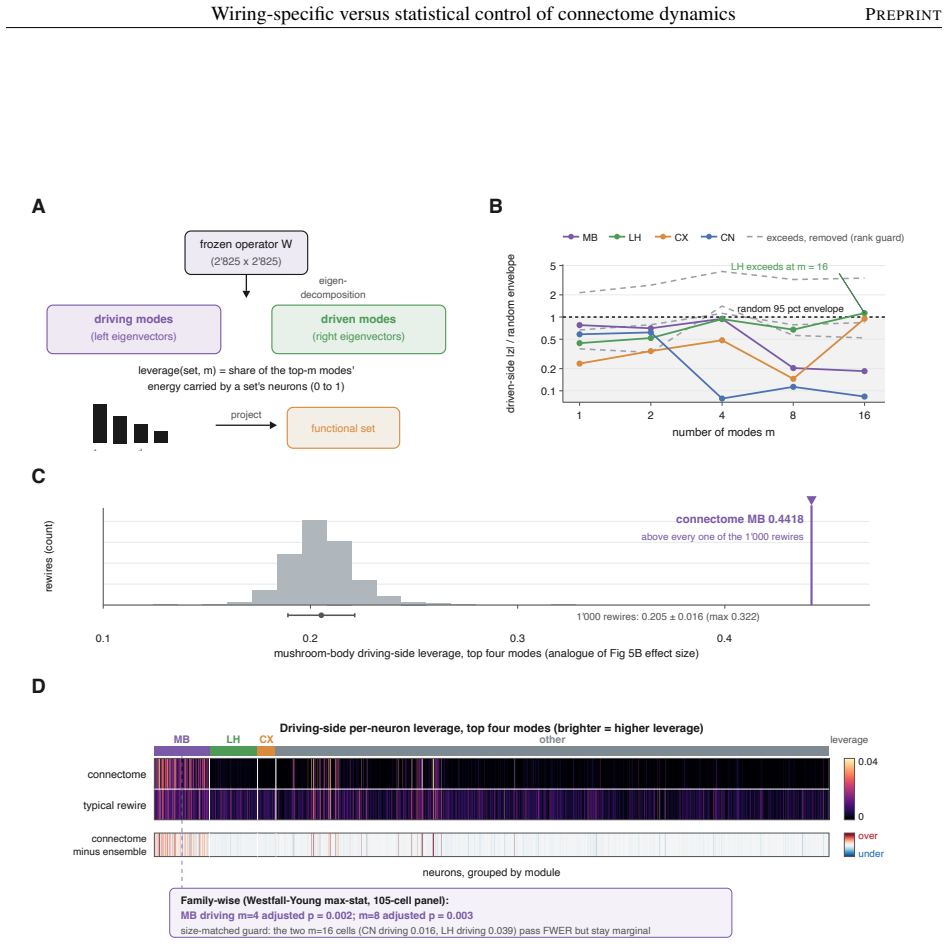
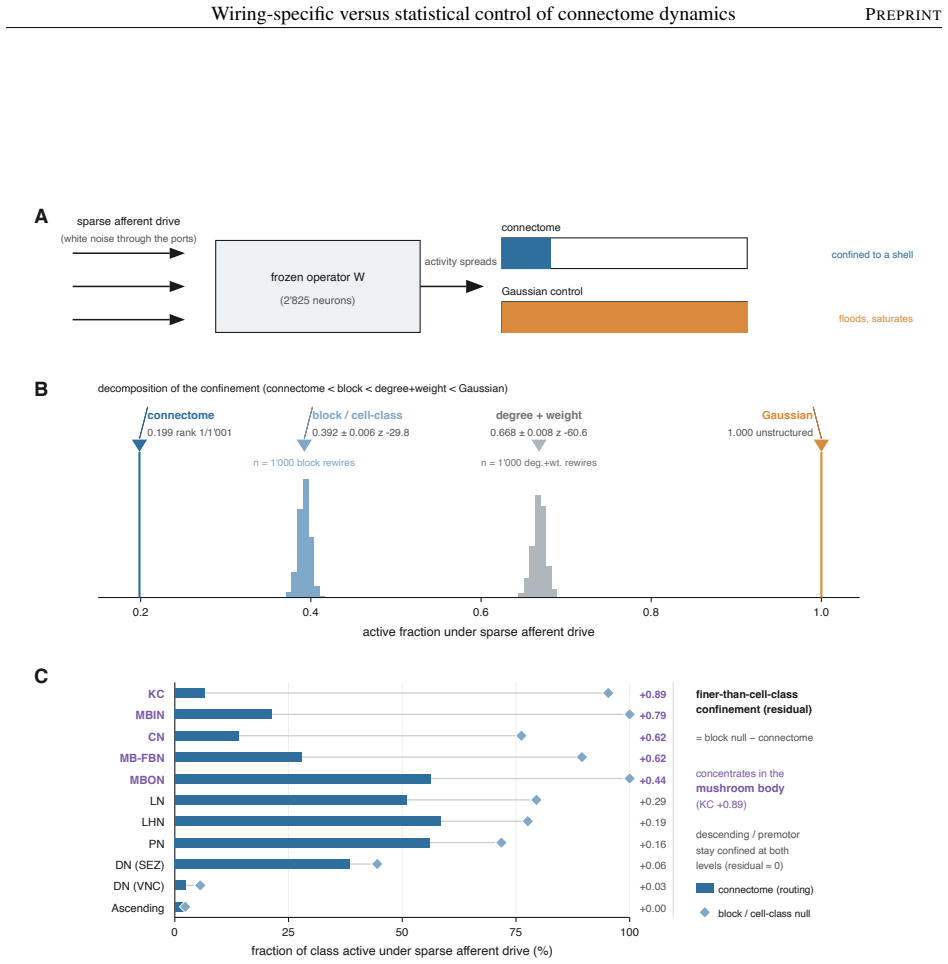
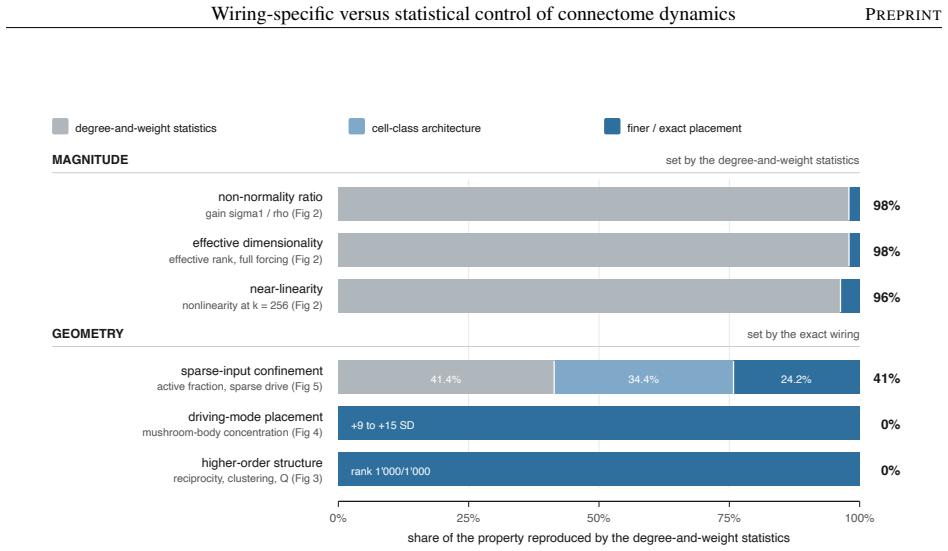
The model's overall dynamical regime, how strongly and how richly it responds, is mostly statistical: networks keeping only the connectome's coarse wiring statistics reproduce it. The wiring beyond these statistics instead sets where activity travels and which circuits shape it. Sparse input is confined to a compact olfactory pathway that randomized networks flood, and the mushroom body, the insect learning center, takes an outsized role in the leading adjoint-side modes, the directions that weigh which neurons shape the recurrent dynamics. Coarse statistics set the regime; the precise pattern of connections sets the geometry.
What carries the argument
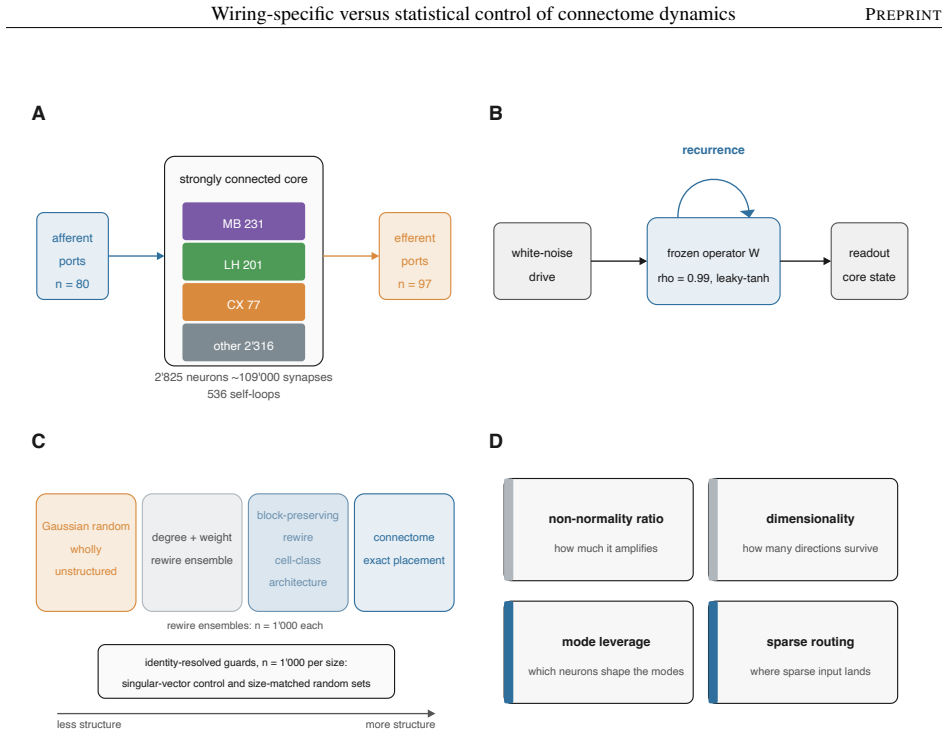
A hierarchy of randomized networks each preserving a coarser description of the wiring, compared against the full connectome inside a fixed rate-based dynamical operator with no fitted single-neuron parameters.
If this is right
- The overall response regime can be reproduced from coarse wiring statistics alone.
- Precise wiring confines sparse inputs to the olfactory pathway rather than allowing them to spread.
- The mushroom body dominates the leading adjoint modes that determine how neurons influence recurrent dynamics.
- Only claims about specific activity paths and circuit roles require the exact connection pattern.
Where Pith is reading between the lines
- The same statistical-versus-specific split could be tested on other reconstructed connectomes to see whether the separation is general.
- Statistical approximations of wiring may suffice for predicting overall response strength but not for circuit-level predictions.
- Experiments that selectively disrupt precise connections could check whether regime strength stays intact while flow geometry changes.
Load-bearing premise
The chosen randomization procedures preserve exactly the intended coarse statistics and do not create artifacts, while the fixed rate-based operator without single-neuron parameters isolates the contribution of precise wiring.
What would settle it
If a different randomization method that still matches the same coarse statistics produces a different dynamical regime, or if direct measurements in the fly brain show response strength that deviates from the statistical prediction while flow patterns match the precise wiring.
Figures
read the original abstract
Electron-microscopy reconstruction now yields complete synaptic wiring diagrams, or connectomes, of entire small brains, including the larval Drosophila, the first insect brain reconstructed in full. How far a wiring diagram alone fixes a circuit's activity, as opposed to the finer physiological detail it does not record, is debated. We run a complete connectome as a fixed, rate-based dynamical operator in which no single-neuron parameter is fitted, so that, at one fixed dynamical regime, the model's behavior reflects the wiring and its connection strengths rather than tuned single-neuron physiology, and compare it against a hierarchy of randomized networks that each preserve a coarser description of the wiring. The model's overall dynamical regime, how strongly and how richly it responds, is mostly statistical: networks keeping only the connectome's coarse wiring statistics reproduce it. The wiring beyond these statistics instead sets where activity travels and which circuits shape it. Sparse input is confined to a compact olfactory pathway that randomized networks flood, and the mushroom body, the insect learning center, takes an outsized role in the leading adjoint-side modes, the directions that weigh which neurons shape the recurrent dynamics. Coarse statistics set the regime; the precise pattern of connections sets the geometry, a separation that clarifies which connectome-based claims rest on wiring alone.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper runs a complete larval Drosophila connectome as a fixed rate-based dynamical operator with no fitted single-neuron parameters. It compares the resulting activity regime and geometry against a hierarchy of randomized networks that successively preserve only coarser wiring statistics. The central claim is that coarse statistics control the overall dynamical regime (response strength and richness), while precise wiring determines activity routing, circuit roles (e.g., mushroom body dominance in adjoint modes), and input confinement.
Significance. If the separation is robust, the work supplies a parameter-free, explicitly controlled framework for attributing dynamical properties to wiring statistics versus specific geometry in a full connectome. This directly addresses a core interpretative question in connectomics and supplies falsifiable predictions about which claims can be made from wiring diagrams alone. The fixed-operator design and adjoint-mode analysis are explicit strengths.
major comments (2)
- [§3] §3 (randomization hierarchy): the claim that the coarsest randomization level fully isolates statistical control requires explicit verification that the procedure does not inadvertently preserve higher-order motifs or degree correlations that could artifactually reproduce the regime; without the precise null-model construction and its validation against the original degree sequence, the attribution of regime properties remains under-constrained.
- [§4.2] §4.2 (adjoint modes): the outsized mushroom-body contribution is reported for the leading modes, but the manuscript does not show that this ranking survives when the same operator is applied to the randomized networks at matched regime strength; without that control the geometry-specific claim is not isolated from the statistical regime.
minor comments (3)
- [Figure 2] Figure 2 legend: the color scale for activity propagation should explicitly state whether it is normalized per network or globally, to allow direct comparison across panels.
- [Methods] Methods, rate-based operator: the single fixed dynamical regime is stated to be held constant, but the precise value of the global gain parameter and its justification should be given in an equation or table.
- [Abstract] The abstract states that sparse input is confined to an olfactory pathway; the corresponding quantitative metric (e.g., fraction of input variance captured) should be defined in the main text.
Simulated Author's Rebuttal
We thank the referee for the constructive comments. We address each major point below and will revise the manuscript to incorporate the requested controls and validations.
read point-by-point responses
-
Referee: [§3] §3 (randomization hierarchy): the claim that the coarsest randomization level fully isolates statistical control requires explicit verification that the procedure does not inadvertently preserve higher-order motifs or degree correlations that could artifactually reproduce the regime; without the precise null-model construction and its validation against the original degree sequence, the attribution of regime properties remains under-constrained.
Authors: We agree that explicit validation of the null-model construction is required to support the attribution. In the revised manuscript we will add a dedicated subsection detailing the randomization algorithm together with quantitative checks confirming that higher-order motifs and degree correlations are not preserved beyond the target coarse statistics. These checks will include motif-count distributions and degree-correlation matrices for the original connectome versus the randomized ensembles. revision: yes
-
Referee: [§4.2] §4.2 (adjoint modes): the outsized mushroom-body contribution is reported for the leading modes, but the manuscript does not show that this ranking survives when the same operator is applied to the randomized networks at matched regime strength; without that control the geometry-specific claim is not isolated from the statistical regime.
Authors: We concur that the geometry-specific claim requires this additional control. The revised manuscript will include the adjoint-mode analysis performed on the randomized networks with dynamical regime strength explicitly matched to the original connectome (via scaling of the effective gain). This will allow direct comparison of mushroom-body participation ranks and thereby isolate the contribution of precise wiring geometry. revision: yes
Circularity Check
No significant circularity
full rationale
The derivation relies on a fixed rate-based operator with no fitted single-neuron parameters, applied uniformly to the connectome and to an explicit hierarchy of randomized controls that preserve successively coarser wiring statistics. The central separation (overall regime set by statistics; geometry and routing set by precise wiring) is obtained by direct comparison of these networks rather than by any self-definitional equation, fitted input renamed as prediction, or load-bearing self-citation. No equation or step reduces to its own inputs by construction; the design is parameter-free and externally falsifiable against the randomization procedure.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption A rate-based dynamical operator with no single-neuron parameters fitted suffices to isolate wiring effects from physiological detail.
Reference graph
Works this paper leans on
-
[1]
URLhttps://www.science.org/doi/abs/10.1126/science.add9330
Michael Winding, Benjamin D. Pedigo, Christopher L. Barnes, Heather G. Patsolic, Youngser Park, Tom Kazimiers, Akira Fushiki, Ingrid V . Andrade, Avinash Khandelwal, Javier Valdes-Aleman, Feng Li, Nadine Randel, Elizabeth Barsotti, Ana Correia, Richard D. Fetter, V olker Hartenstein, Carey E. Priebe, Joshua T. V ogelstein, Albert Cardona, and Marta Zlatic...
-
[2]
S. J. Cook, T. A. Jarrell, C. A. Brittin, Y . Wang, A. E. Bloniarz, M. A. Yakovlev, K. C. Q. Nguyen, L. T.-H. Tang, E. A. Bayer, J. S. Duerr, H. E. Bülow, O. Hobert, D. H. Hall, and S. W. Emmons. Whole-animal connectomes of both Caenorhabditis elegans sexes.Nature, 571(7763):63–71, 2019. doi: 10.1038/s41586-019-1352-7
-
[3]
Sterling, Philipp Schlegel, et al
Sven Dorkenwald, Arie Matsliah, Amy R. Sterling, Philipp Schlegel, et al. Neuronal wiring diagram of an adult brain.Nature, 634(8032):124–138, 2024. doi: 10.1038/s41586-024-07558-y
-
[4]
D. G. C. Hildebrand, M. Cicconet, R. M. Torres, et al. Whole-brain serial-section electron microscopy in larval zebrafish.Nature, 545(7654):345–349, 2017. doi: 10.1038/nature22356
-
[5]
Functional connectomics spanning multiple areas of mouse visual cortex.Nature,
The MICrONS Consortium. Functional connectomics spanning multiple areas of mouse visual cortex.Nature,
-
[6]
doi: 10.1038/s41586-025-08790-w
-
[7]
Alexander Shapson-Coe, Michał Januszewski, Daniel R. Berger, et al. A petavoxel fragment of human cerebral cortex reconstructed at nanoscale resolution.Science, 384(6696):eadk4858, 2024. doi: 10.1126/science.adk4858
-
[8]
Manuel Beiran and Ashok Litwin-Kumar. Prediction of neural activity in connectome-constrained recurrent networks.Nature Neuroscience, 28:2561–2574, 2025. doi: 10.1038/s41593-025-02080-4
-
[9]
Katharina Eichler, Feng Li, Ashok Litwin-Kumar, Youngser Park, Ingrid V . Andrade, Casey M. Schneider-Mizell, Timo Saumweber, Arnim Huser, Claire Eschbach, Bertram Gerber, Richard D. Fetter, James W. Truman, Carey E. Priebe, L. F. Abbott, Andreas S. Thum, Marta Zlatic, and Albert Cardona. The complete connectome of a learning and memory centre in an insec...
-
[10]
Berck, Avinash Khandelwal, Lindsey Claus, Luis Hernandez-Nunez, Guangwei Si, Christopher J
Matthew E. Berck, Avinash Khandelwal, Lindsey Claus, Luis Hernandez-Nunez, Guangwei Si, Christopher J. Tabone, Feng Li, James W. Truman, Richard D. Fetter, Matthieu Louis, Aravinthan D. T. Samuel, and Albert Cardona. The wiring diagram of a glomerular olfactory system.eLife, 5:e14859, 2016. doi: 10.7554/eLife.14859
-
[11]
Schneider-Mizell, Mei Shao, Rebecca Arruda, Katharina Eichler, Javier Valdes-Aleman, Tomoko Ohyama, Andreas S
Claire Eschbach, Akira Fushiki, Michael Winding, Casey M. Schneider-Mizell, Mei Shao, Rebecca Arruda, Katharina Eichler, Javier Valdes-Aleman, Tomoko Ohyama, Andreas S. Thum, Bertram Gerber, Richard D. Fetter, James W. Truman, Ashok Litwin-Kumar, Albert Cardona, and Marta Zlatic. Recurrent architecture for adaptive regulation of learning in the insect bra...
2020
-
[12]
The central complex of the larval fruit fly brain.bioRxiv, 2025
Laura Lungu, Nicolò Ceffa, Michael Clayton, Marta Zlatic, and Albert Cardona. The central complex of the larval fruit fly brain.bioRxiv, 2025. doi: 10.1101/2025.09.30.679510. Preprint
-
[13]
L. E. Suárez, A. Mihalik, F. Milisav, K. Marshall, M. Li, P. E. Vértes, G. Lajoie, and B. Misic. Connectome- based reservoir computing with the conn2res toolbox.Nature Communications, 15, 2024. doi: 10.1038/ s41467-024-44900-4
2024
-
[14]
L. Costi, A. Hadjiivanov, D. Dold, Z. F. Hale, and D. Izzo. The Drosophila connectome as a computational reservoir for time-series prediction.Biomimetics, 10(5):341, 2025. doi: 10.3390/biomimetics10050341
-
[15]
S. Yu, Z. Qin, T. Liu, B. Xu, R. J. V ogelstein, J. Brown, and J. T. V ogelstein. Biological processing units: leveraging an insect connectome to pioneer biofidelic neural architectures. InArtificial General Intelligence (AGI 2025), volume 16058 ofLNCS. Springer, 2025. doi: 10.1007/978-3-032-00800-8\_32. arXiv:2507.10951
-
[16]
Specificity and stability in topology of protein networks.Science, 296(5569): 910–913, 2002
Sergei Maslov and Kim Sneppen. Specificity and stability in topology of protein networks.Science, 296(5569): 910–913, 2002. doi: 10.1126/science.1065103
-
[17]
Horn and Charles R
Roger A. Horn and Charles R. Johnson.Matrix Analysis. Cambridge University Press, Cambridge, 2 edition,
-
[18]
doi: 10.1017/CBO9781139020411
-
[19]
Masuda-Nakagawa, Kei Ito, Takeshi Awasaki, and Cahir J
Liria M. Masuda-Nakagawa, Kei Ito, Takeshi Awasaki, and Cahir J. O’Kane. A single GABAergic neuron mediates feedback of odor-evoked signals in the mushroom body of larval Drosophila.Frontiers in Neural Circuits, 8:35, 2014. doi: 10.3389/fncir.2014.00035
-
[20]
Staba, Fabio Ferrarelli, Chiara Cirelli, Giulio Tononi, and Itzhak Fried
Nino Mancini, Juliane Thoener, Esmeralda Tafani, Dennis Pauls, Oded Mayseless, Martin Strauch, Katharina Eichler, Andrew Champion, Oliver Kobler, Denise Weber, Edanur Sen, Ali ´ce Weiglein, V olker Hartenstein, Charalampos-Chrysovalantis Chytoudis-Peroudis, Tihana Jovanic, Andreas S. Thum, Astrid Rohwedder, Michael Schleyer, and Bertram Gerber. Rewarding ...
-
[21]
R. Betzel, M. G. Puxeddu, and C. Seguin. Hierarchical communities in the larval Drosophila connectome: links to cellular annotations and network topology.Proceedings of the National Academy of Sciences, 121(38): e2320177121, 2024. doi: 10.1073/pnas.2320177121
-
[22]
Turner, Maxim Bazhenov, and Gilles Laurent
Glenn C. Turner, Maxim Bazhenov, and Gilles Laurent. Olfactory representations by Drosophila mushroom body neurons.Journal of Neurophysiology, 99:734–746, 2008. doi: 10.1152/jn.01283.2007
-
[23]
Kyle S. Honegger, Robert A. A. Campbell, and Glenn C. Turner. Cellular-resolution population imaging reveals robust sparse coding in the Drosophila mushroom body.Journal of Neuroscience, 31(33):11772–11785, 2011. doi: 10.1523/JNEUROSCI.1099-11.2011
-
[24]
Ashok Litwin-Kumar, Kameron Decker Harris, Richard Axel, Haim Sompolinsky, and L. F. Abbott. Optimal degrees of synaptic connectivity.Neuron, 93(5):1153–1164.e7, 2017. doi: 10.1016/j.neuron.2017.01.030
-
[25]
D. G. Clark, O. Marschall, A. van Meegen, and A. Litwin-Kumar. Connectivity structure and dynamics of nonlinear recurrent neural networks.Physical Review X, 15(4):041019, 2025. doi: 10.1103/2jt7-c8cq. arXiv:2409.01969
-
[26]
P. Yadav, P. Shinde, and A. Singh. Brain rewiring during development: a comparative analysis of larval and adult Drosophila melanogaster connectomes.Network Neuroscience, 9(4):1299–1322, 2025. doi: 10.1162/NETN.a.26
-
[27]
C. Seguin, M. G. Puxeddu, J. Faskowitz, R. F. Betzel, and O. Sporns. Connectome architecture favours within- module diffusion and between-module routing.bioRxiv, 2025. doi: 10.1101/2025.02.10.637586. Preprint, posted 2025-02-11. 20
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.