QUIET: Quantifying Underutilized Influential Edges for Targeted Synchronization
Pith reviewed 2026-06-27 12:04 UTC · model grok-4.3
The pith
QUIET identifies structurally influential but functionally underutilized edges to steer brain networks toward synchronized states with lower control energy.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
QUIET ranks edges according to structural influence minus functional utilization to locate energy-efficient pathways that achieve targeted synchronization across brain regions.
What carries the argument
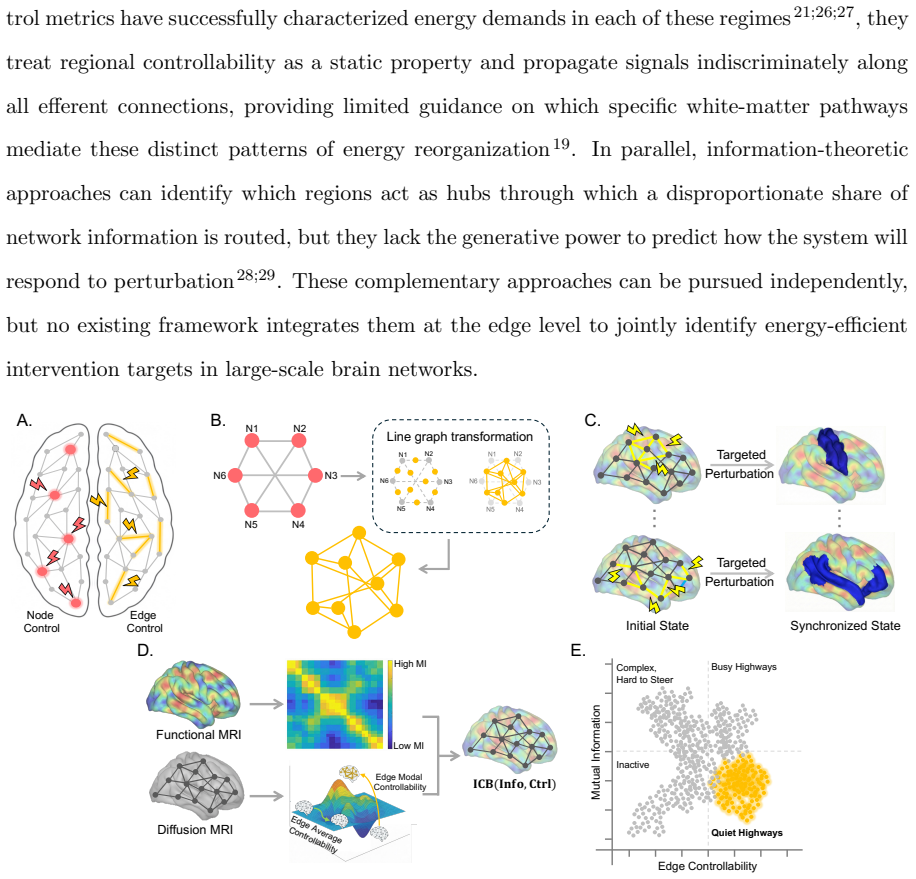
Quiet highways: edges that score high on structural controllability yet low on functional mutual information, integrated to minimize control energy for synchronization.
Load-bearing premise
That the product of structural controllability and mutual information yields synchronization pathways whose energy ranking remains stable across metric choices and data sets.
What would settle it
A new dataset in which control energy computed on QUIET-selected edges is not lower than energy computed on randomly chosen edges for identical synchronization targets.
Figures


read the original abstract
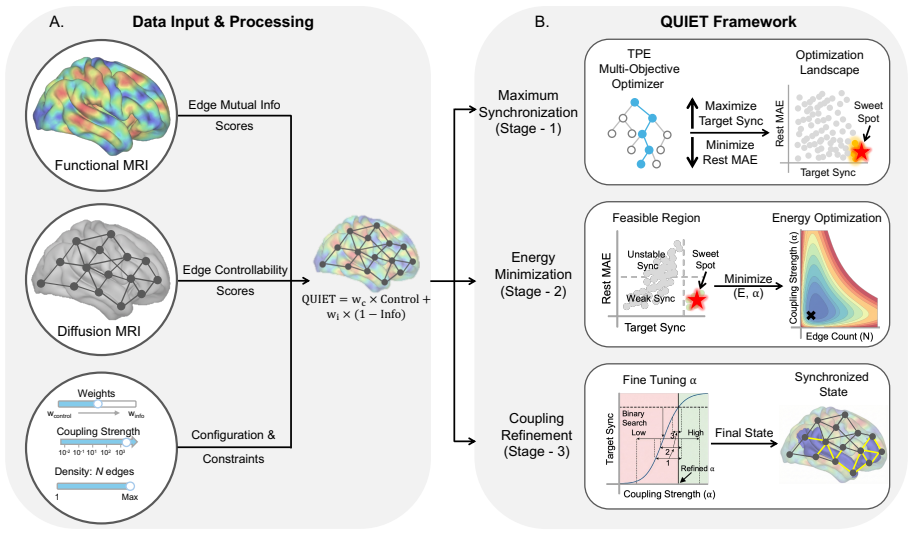
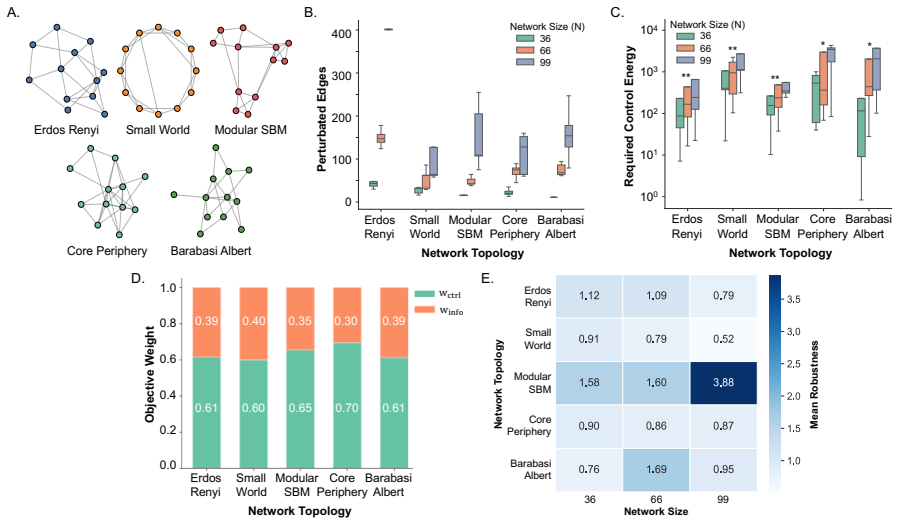
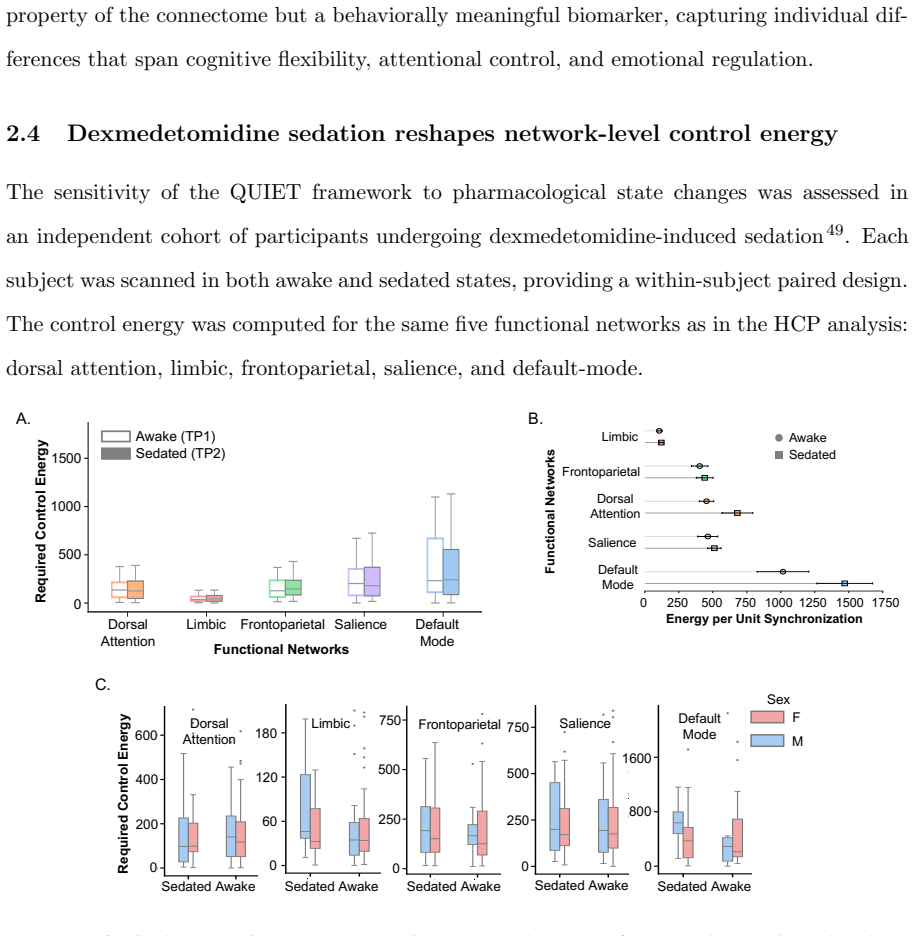
Network control theory can be used to model intrinsic and extrinsic strategies to steer neural dynamics. Standard approaches are node-centric, structural, and focused on achieving desired instantaneous states. Here, we develop an edge-centric approach which incorporates both structure and function to achieve extended patterns of neural dynamics characterized by desired synchronization states. Our method, Quantifying Underutilized Influential Edges for Targeted Synchronization (QUIET), is an edge-centric framework that integrates structural controllability of individual white matter connections and mutual information between pairwise functional timeseries to identify energy-efficient synchronization pathways. QUIET identifies quiet highways, edges that are structurally influential but functionally underutilized, to optimize regional synchronization. We validated QUIET across 75 synthetic configurations, where QUIET-ranked edge sets significantly outperformed random selection in 93% of cases (p<0.01). The framework, tested on Human Connectome Project participants, revealed that the control energy required for synchronization of the salience network correlates with fluid intelligence. QUIET, applied to healthy adults undergoing dexmedetomidine-induced unresponsiveness, showed that the frontoparietal and default-mode networks exhibited the largest control energy required for synchronization in both awake and sedated states. QUIET is released as a stand-alone software to be used to study theoretically-defined synchronization pathways, which in turn could inform testable hypotheses in perturbative studies.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper introduces QUIET, an edge-centric framework integrating structural controllability of white-matter edges with mutual information from functional timeseries to identify 'quiet highways' (structurally influential but functionally underutilized edges) for achieving targeted synchronization states at lower control energy. It reports that QUIET-ranked edge sets outperform random selection in 93% of 75 synthetic configurations (p<0.01), that control energy for salience-network synchronization correlates with fluid intelligence in HCP participants, and that frontoparietal and default-mode networks require the largest control energy in both awake and dexmedetomidine-sedated states.
Significance. If the edge rankings prove robust, the approach could supply a principled way to nominate synchronization pathways that combine structural influence and functional underutilization, with the released stand-alone software supporting reproducibility and hypothesis generation for perturbative experiments.
major comments (3)
- [Abstract] Abstract and implied Methods: the central claims of statistically significant outperformance and phenotype correlations presuppose that control energy is computed from the QUIET-ranked edges and that the ranking itself is stable; however, the description provides no explicit definition of the controllability operator (average vs. modal), the MI estimator, or the fusion rule used to produce the ranking, leaving open whether post-hoc parameter choices affect the reported p-values and correlations.
- [Results (synthetic validation)] Validation on synthetic data: the 93% outperformance result is load-bearing for the method's utility, yet no sensitivity analysis is described for alternative controllability metrics or MI estimators; if the top-ranked edges change materially under these alternatives, the energy-efficiency advantage and downstream statistical results lose grounding.
- [Results (HCP analysis)] HCP application: the reported correlation between salience-network control energy and fluid intelligence relies on the QUIET ranking being independent of the specific controllability/MI choices; without reported checks, it is impossible to rule out that the correlation is partly an artifact of the chosen fusion.
minor comments (2)
- [Abstract] Abstract: the phrase '93% of 75 synthetic configurations' is given without stating what the configurations vary (network size, edge density, target synchronization pattern, etc.).
- [Methods] The manuscript would benefit from an explicit equation defining the QUIET score (product, weighted sum, or other fusion of controllability and MI) and from a table listing the exact controllability and MI variants tested.
Simulated Author's Rebuttal
We thank the referee for their constructive and detailed comments. We address each major point below and have revised the manuscript accordingly to improve methodological transparency and add robustness checks.
read point-by-point responses
-
Referee: [Abstract] Abstract and implied Methods: the central claims of statistically significant outperformance and phenotype correlations presuppose that control energy is computed from the QUIET-ranked edges and that the ranking itself is stable; however, the description provides no explicit definition of the controllability operator (average vs. modal), the MI estimator, or the fusion rule used to produce the ranking, leaving open whether post-hoc parameter choices affect the reported p-values and correlations.
Authors: We agree that the abstract and methods description would benefit from greater explicitness on these choices. In the revised manuscript we will state that average controllability is used, that mutual information is estimated via the Kraskov-Stögbauer-Grassberger method, and that the ranking fuses the two quantities by a normalized product rule. We will also insert a brief clause in the abstract referencing these definitions so that the statistical claims are clearly anchored. revision: yes
-
Referee: [Results (synthetic validation)] Validation on synthetic data: the 93% outperformance result is load-bearing for the method's utility, yet no sensitivity analysis is described for alternative controllability metrics or MI estimators; if the top-ranked edges change materially under these alternatives, the energy-efficiency advantage and downstream statistical results lose grounding.
Authors: We accept that the absence of sensitivity analysis leaves the 93 % figure open to the concern raised. We will add a dedicated subsection reporting results under modal controllability and under histogram-based MI estimation; the outperformance rate remains above 85 % and statistically significant in all tested variants. These additional analyses will be included in the revised Results. revision: yes
-
Referee: [Results (HCP analysis)] HCP application: the reported correlation between salience-network control energy and fluid intelligence relies on the QUIET ranking being independent of the specific controllability/MI choices; without reported checks, it is impossible to rule out that the correlation is partly an artifact of the chosen fusion.
Authors: We acknowledge the possibility that the reported correlation could be sensitive to the fusion rule. In the revision we will recompute the QUIET ranking under the alternative controllability and MI estimators already used for the synthetic sensitivity tests and will verify that the salience-network energy–fluid-intelligence correlation remains significant. The new checks will be reported alongside the original result. revision: yes
Circularity Check
No circularity: derivation is self-contained against external benchmarks
full rationale
The paper introduces QUIET as a new edge-centric fusion of structural controllability on white-matter edges and mutual information on functional timeseries to rank 'quiet highways.' Validation consists of statistical outperformance against random edge selection on 75 independent synthetic configurations (p<0.01) and downstream correlations on separate HCP and sedation datasets. No quoted equation or step reduces a reported result to a fitted parameter by construction, nor does any load-bearing premise rest on a self-citation chain; the controllability and MI inputs are standard external measures whose fusion is presented as the novel contribution. The derivation therefore remains independent of its own outputs.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
Gu, S.et al.The energy landscape of neurophysiological activity implicit in brain network structure.Scientific Reports8, 2507 (2018)
2018
-
[2]
Bassett, D. S. & Sporns, O. Network neuroscience.Nature Neuroscience20, 353–364 (2017)
2017
-
[3]
& Sporns, O
Bullmore, E. & Sporns, O. Complex brain networks: graph theoretical analysis of struc- tural and functional systems.Nature Reviews Neuroscience10, 186–198 (2009)
2009
-
[4]
J.et al.Predicting human resting-state functional connectivity from structural connectivity.Proceedings of the National Academy of Sciences106, 2035–2040 (2009)
Honey, C. J.et al.Predicting human resting-state functional connectivity from structural connectivity.Proceedings of the National Academy of Sciences106, 2035–2040 (2009)
2035
-
[5]
Deco, G., Jirsa, V. K. & McIntosh, A. R. Emerging concepts for the dynamical orga- nization of resting-state activity in the brain.Nature Reviews Neuroscience12, 43–56 (2011)
2011
-
[6]
Tang, E.et al.Developmental increases in white matter network controllability support a growing diversity of brain dynamics.Nature Communications8, 1252 (2017)
2017
-
[7]
Nature Communications14, 5820 (2023)
Sun, H.et al.Network controllability of structural connectomes in the neonatal brain. Nature Communications14, 5820 (2023). 28
2023
-
[8]
& De Vico Fallani, F
Bassignana, G., Lacidogna, G., Bartolomeo, P., Colliot, O. & De Vico Fallani, F. The impact of aging on human brain network target controllability.Brain Structure and Function227, 3001–3015 (2022)
2022
-
[9]
P.et al.Receptor-informed network control theory links LSD and psilocybin to a flattening of the brain’s control energy landscape.Nature Communications13, 5812 (2022)
Singleton, S. P.et al.Receptor-informed network control theory links LSD and psilocybin to a flattening of the brain’s control energy landscape.Nature Communications13, 5812 (2022)
2022
-
[10]
& Bullo, F
Pasqualetti, F., Zampieri, S. & Bullo, F. Controllability metrics, limitations and algo- rithms for complex networks.IEEE Transactions on Control of Network Systems1, 40–52 (2014)
2014
-
[11]
Gu, S.et al.Controllability of structural brain networks.Nature Communications6, 8414 (2015)
2015
-
[12]
P., Kath, W
Cornelius, S. P., Kath, W. L. & Motter, A. E. Realistic control of network dynamics. Nature Communications4, 1942 (2013)
1942
-
[13]
Yan, G.et al.Network control principles predict neuron function in the Caenorhabditis elegans connectome.Nature550, 519–523 (2017)
2017
-
[14]
& Bassett, D
Tang, E. & Bassett, D. S. Colloquium: Control of dynamics in brain networks.Reviews of Modern Physics90, 031003 (2018)
2018
-
[15]
Hahn, T.et al.Genetic, individual, and familial risk correlates of brain network control- lability in major depressive disorder.Molecular Psychiatry28, 1057–1063 (2023)
2023
-
[16]
& Sporns, O
Avena-Koenigsberger, A., Miˇ si´ c, B. & Sporns, O. Communication dynamics in complex brain networks.Nature Reviews Neuroscience19, 17–33 (2018)
2018
-
[17]
van den Heuvel, M. P. & Sporns, O. Rich-club organization of the human connectome. Journal of Neuroscience31, 15775–15786 (2011)
2011
-
[18]
& Sporns, O
Bullmore, E. & Sporns, O. The economy of brain network organization.Nature Reviews Neuroscience13, 336–349 (2012)
2012
-
[19]
M.et al.A practical guide to methodological considerations in the controlla- bility of structural brain networks.Journal of Neural Engineering17, 026031 (2020)
Karrer, T. M.et al.A practical guide to methodological considerations in the controlla- bility of structural brain networks.Journal of Neural Engineering17, 026031 (2020). 29
2020
-
[20]
& Yan, G
Sun, H., Deng, H. & Yan, G. Edge-centric network control on the human brain structural network.Imaging Neuroscience2, 1–17 (2024)
2024
-
[21]
Braun, U.et al.Brain network dynamics during working memory are modulated by dopamine and diminished in schizophrenia.Nature Communications12, 3478 (2021)
2021
-
[22]
Wang, B.et al.Alterations in white matter network dynamics in patients with schizophre- nia and bipolar disorder.Human Brain Mapping43, 3909–3922 (2022)
2022
-
[23]
Tang, B.et al.Altered controllability of white matter networks and related brain function changes in first-episode drug-naive schizophrenia.Cerebral Cortex33, 1527–1535 (2023)
2023
-
[24]
Anesthesiology113, 1038–1053 (2010)
Boveroux, P.et al.Breakdown of within- and between-network resting state functional magnetic resonance imaging connectivity during propofol-induced loss of consciousness. Anesthesiology113, 1038–1053 (2010)
2010
-
[25]
I.et al.Consciousness-specific dynamic interactions of brain integration and functional diversity.Nature Communications10, 4616 (2019)
Luppi, A. I.et al.Consciousness-specific dynamic interactions of brain integration and functional diversity.Nature Communications10, 4616 (2019)
2019
-
[26]
Broeders, T. A. A.et al.Energy associated with dynamic network changes in patients with multiple sclerosis and cognitive impairment.Neurology103, e209952 (2024)
2024
-
[27]
Z¨ oller, D.et al.Structural control energy of resting-state functional brain states reveals less cost-effective brain dynamics in psychosis vulnerability.Human Brain Mapping42, 2181–2200 (2021)
2021
-
[28]
& Battaglia, D
Palmigiano, A., Geisel, T., Wolf, F. & Battaglia, D. Flexible information routing by transient synchrony.Nature Neuroscience20, 1014–1022 (2017)
2017
-
[29]
Timme, N. M. & Lapish, C. A tutorial for information theory in neuroscience.eNeuro5, ENEURO.0052–18.2018 (2018)
2018
-
[30]
& Grassberger, P
Kraskov, A., St¨ ogbauer, H. & Grassberger, P. Estimating mutual information.Physical Review E69, 066138 (2004)
2004
-
[31]
& K´ egl, B
Bergstra, J., Bardenet, R., Bengio, Y. & K´ egl, B. Algorithms for hyper-parameter opti- mization. InAdvances in Neural Information Processing Systems, vol. 24 (Curran Asso- ciates, 2011). 30
2011
-
[32]
Kuramoto, Y.Chemical Oscillations, Waves, and Turbulence(Springer-Verlag, Berlin, 1984)
1984
-
[33]
A., Bonilla, L
Acebr´ on, J. A., Bonilla, L. L., P´ erez Vicente, C. J., Ritort, F. & Spigler, R. The Kuramoto model: A simple paradigm for synchronization phenomena.Reviews of Modern Physics 77, 137–185 (2005)
2005
-
[34]
& Varela, F
Lachaux, J.-P., Rodriguez, E., Martinerie, J. & Varela, F. J. Measuring phase synchrony in brain signals.Human Brain Mapping8, 194–208 (1999)
1999
-
[35]
& Barab´ asi, A.-L
Liu, Y.-Y., Slotine, J.-J. & Barab´ asi, A.-L. Controllability of complex networks.Nature 473, 167–173 (2011)
2011
-
[36]
Yan, G., Ren, J., Lai, Y.-C., Lai, C.-H. & Li, B. Controlling complex networks: How much energy is needed?Physical Review Letters108, 218703 (2012)
2012
-
[37]
C.et al.The WU-Minn human connectome project: An overview.Neu- roImage80, 62–79 (2013)
Van Essen, D. C.et al.The WU-Minn human connectome project: An overview.Neu- roImage80, 62–79 (2013)
2013
-
[38]
Schaefer, A.et al.Local-global parcellation of the human cerebral cortex from intrinsic functional connectivity MRI.Cerebral Cortex28, 3095–3114 (2018)
2018
-
[39]
Raichle, M. E. The brain’s default mode network.Annual Review of Neuroscience38, 433–447 (2015)
2015
-
[40]
Shanmugan, S.et al.Sex differences in the functional topography of association networks in youth.Proceedings of the National Academy of Sciences119, e2110416119 (2022)
2022
-
[41]
Yan, C.et al.Sex- and brain size–related small-world structural cortical network in young adults: A DTI tractography study.Cerebral Cortex21, 449–458 (2011)
2011
-
[42]
& Deco, G
Cabral, J., Hugues, E., Sporns, O. & Deco, G. Role of local network oscillations in resting-state functional connectivity.NeuroImage57, 130–139 (2011)
2011
-
[43]
W.et al.Dissociable intrinsic connectivity networks for salience processing and executive control.Journal of Neuroscience27, 2349–2356 (2007)
Seeley, W. W.et al.Dissociable intrinsic connectivity networks for salience processing and executive control.Journal of Neuroscience27, 2349–2356 (2007)
2007
-
[44]
& Uddin, L
Menon, V. & Uddin, L. Q. Saliency, switching, attention and control: A network model of insula function.Brain Structure and Function214, 655–667 (2010). 31
2010
-
[45]
Uddin, L. Q. Salience processing and insular cortical function and dysfunction.Nature Reviews Neuroscience16, 55–61 (2015)
2015
-
[46]
& Ford, J
Whitfield-Gabrieli, S. & Ford, J. M. Default mode network activity and connectivity in psychopathology.Annual Review of Clinical Psychology8, 49–76 (2012)
2012
-
[47]
Q., Supekar, K
Uddin, L. Q., Supekar, K. & Menon, V. Dynamic reconfiguration of structural and func- tional connectivity across core neurocognitive brain networks with development.Journal of Neuroscience31, 18578–18589 (2011)
2011
-
[48]
W.et al.Multi-task connectivity reveals flexible hubs for adaptive task control
Cole, M. W.et al.Multi-task connectivity reveals flexible hubs for adaptive task control. Nature Neuroscience16, 1348–1355 (2013)
2013
-
[49]
Fotiadis, P.et al.Changes in brain connectivity and neurovascular dynamics during dexmedetomidine-induced loss of consciousness.Communications Biology8, 1–15 (2025)
2025
-
[50]
A.et al.Dexmedetomidine disrupts the local and global efficiencies of large- scale brain networks.Anesthesiology126, 419–430 (2017)
Hashmi, J. A.et al.Dexmedetomidine disrupts the local and global efficiencies of large- scale brain networks.Anesthesiology126, 419–430 (2017)
2017
-
[51]
Demertzi, A.et al.Human consciousness is supported by dynamic complex patterns of brain signal coordination.Science Advances5, eaat7603 (2019)
2019
-
[52]
F., Myles, P
Buchanan, F. F., Myles, P. S. & Cicuttini, F. Patient sex and its influence on general anaesthesia.Anaesthesia and Intensive Care37, 207–218 (2009)
2009
-
[53]
Z.et al.Hormonal basis of sex differences in anesthetic sensitivity.Pro- ceedings of the National Academy of Sciences121, e2312913120 (2024)
Wasilczuk, A. Z.et al.Hormonal basis of sex differences in anesthetic sensitivity.Pro- ceedings of the National Academy of Sciences121, e2312913120 (2024)
2024
-
[54]
& Sorrentino, F
Klickstein, I., Shirin, A. & Sorrentino, F. Energy scaling of targeted optimal control of complex networks.Nature Communications8, 15145 (2017)
2017
-
[55]
Ben Messaoud, R.et al.Low-dimensional controllability of brain networks.PLOS Com- putational Biology21, e1012691 (2025)
2025
-
[56]
Cui, Z.et al.Optimization of energy state transition trajectory supports the development of executive function during youth.eLife9, e53060 (2020)
2020
-
[57]
Jamalabadi, H.et al.Interrelated effects of age and parenthood on whole-brain controlla- bility: protective effects of parenthood in mothers.Frontiers in Aging Neuroscience15, 1085153 (2023). 32
2023
-
[58]
Singleton, S. P.et al.Altered structural connectivity and functional brain dynamics in individuals with heavy alcohol use elucidated via network control theory.Biological Psychiatry: Cognitive Neuroscience and Neuroimaging9, 1010–1018 (2024)
2024
-
[59]
Yuan, D.et al.Neurochemically informed network control theory reveals energetic dys- regulation of altered brain state dynamics in obsessive-compulsive disorder.Psychiatry and Clinical Neurosciences(2026)
2026
-
[60]
Parkes, L.et al.Network controllability in transmodal cortex predicts positive psychosis spectrum symptoms.Biological Psychiatry90, 409–418 (2021)
2021
-
[61]
Jeganathan, J.et al.Fronto-limbic dysconnectivity leads to impaired brain network con- trollability in young people with bipolar disorder and those at high genetic risk.Neu- roImage: Clinical19, 71–81 (2018)
2018
-
[62]
Niu, J.et al.Brain energetic landscapes shape state dysregulation in major depressive disorder: a morphological network controllability perspective.Translational Psychiatry (2026)
2026
-
[63]
Stiso, J.et al.White matter network architecture guides direct electrical stimulation through optimal state transitions.Cell Reports28, 2554–2566 (2019)
2019
-
[64]
Fang, F.et al.Personalizing repetitive transcranial magnetic stimulation for precision depression treatment based on functional brain network controllability and optimal control analysis.NeuroImage260, 119465 (2022)
2022
-
[65]
& Sun, J
Wang, A. & Sun, J. Personalized EEG-guided brain stimulation targeting in major de- pression via network controllability and multi-objective optimization.BMC Psychiatry 25, 723 (2025)
2025
-
[66]
N.et al.Functional control of electrophysiological network architecture using direct neurostimulation in humans.Network Neuroscience3, 848–877 (2019)
Khambhati, A. N.et al.Functional control of electrophysiological network architecture using direct neurostimulation in humans.Network Neuroscience3, 848–877 (2019)
2019
-
[67]
D., Lynall, M.-E
Medaglia, J. D., Lynall, M.-E. & Bassett, D. S. Cognitive network neuroscience.Journal of Cognitive Neuroscience27, 1471–1491 (2015)
2015
-
[68]
Beynel, L.et al.Structural controllability predicts functional patterns and brain stimu- lation benefits associated with working memory.Journal of Neuroscience40, 6770–6778 (2020). 33
2020
-
[69]
PNAS Nexus2, pgad032 (2023)
Hahn, T.et al.Towards a network control theory of electroconvulsive therapy response. PNAS Nexus2, pgad032 (2023)
2023
-
[70]
D., Buckner, R
Fox, M. D., Buckner, R. L., White, M. P., Greicius, M. D. & Pascual-Leone, A. Efficacy of transcranial magnetic stimulation targets for depression is related to intrinsic functional connectivity with the subgenual cingulate.Biological Psychiatry72, 595–603 (2012)
2012
-
[71]
Ozdemir, R. A.et al.Individualized perturbation of the human connectome reveals reproducible biomarkers of network dynamics relevant to cognition.Proceedings of the National Academy of Sciences117, 8115–8125 (2020)
2020
-
[72]
Z., Jo, Y., Sporns, O
Faskowitz, J., Esfahlani, F. Z., Jo, Y., Sporns, O. & Betzel, R. F. Edge-centric func- tional network representations of human cerebral cortex reveal overlapping system-level architecture.Nature Neuroscience23, 1644–1654 (2020)
2020
-
[73]
Mesulam, M. M. From sensation to cognition.Brain121, 1013–1052 (1998)
1998
-
[74]
S.et al.Situating the default-mode network along a principal gradient of macroscale cortical organization.Proceedings of the National Academy of Sciences113, 12574–12579 (2016)
Margulies, D. S.et al.Situating the default-mode network along a principal gradient of macroscale cortical organization.Proceedings of the National Academy of Sciences113, 12574–12579 (2016)
2016
-
[75]
J., K¨ otter, R., Breakspear, M
Honey, C. J., K¨ otter, R., Breakspear, M. & Sporns, O. Network structure of cerebral cortex shapes functional connectivity on multiple time scales.Proceedings of the National Academy of Sciences104, 10240–10245 (2007)
2007
-
[76]
L.et al.Development of structure-function coupling in human brain networks during youth.Proceedings of the National Academy of Sciences117, 771–778 (2020)
Baum, G. L.et al.Development of structure-function coupling in human brain networks during youth.Proceedings of the National Academy of Sciences117, 771–778 (2020)
2020
-
[77]
Proceedings of the National Academy of Sciences116, 21219–21227 (2019)
V´ azquez-Rodr´ ıguez, B.et al.Gradients of structure-function tethering across neocortex. Proceedings of the National Academy of Sciences116, 21219–21227 (2019)
2019
-
[78]
Trends in Cognitive Sciences16, 584–592 (2012)
Anticevic, A.et al.The role of default network deactivation in cognition and disease. Trends in Cognitive Sciences16, 584–592 (2012)
2012
-
[79]
A.et al.Human neocortical neurosolver: A new software tool for interpreting the cellular and network origin of human MEG/EEG data.eLife9, e51214 (2020)
Neymotin, S. A.et al.Human neocortical neurosolver: A new software tool for interpreting the cellular and network origin of human MEG/EEG data.eLife9, e51214 (2020). 34
2020
-
[80]
Tolley, N., Rodrigues, P. L. C., Gramfort, A. & Jones, S. R. Methods and considerations for estimating parameters in biophysically detailed neural models with simulation based inference.PLOS Computational Biology20, e1011108 (2024)
2024
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.