Mechanistic mathematical model of the in vitro infection dynamics of Bunyamwera and Batai viruses including MOI-dependent shortening of the eclipse phase
Pith reviewed 2026-06-26 12:38 UTC · model grok-4.3
The pith
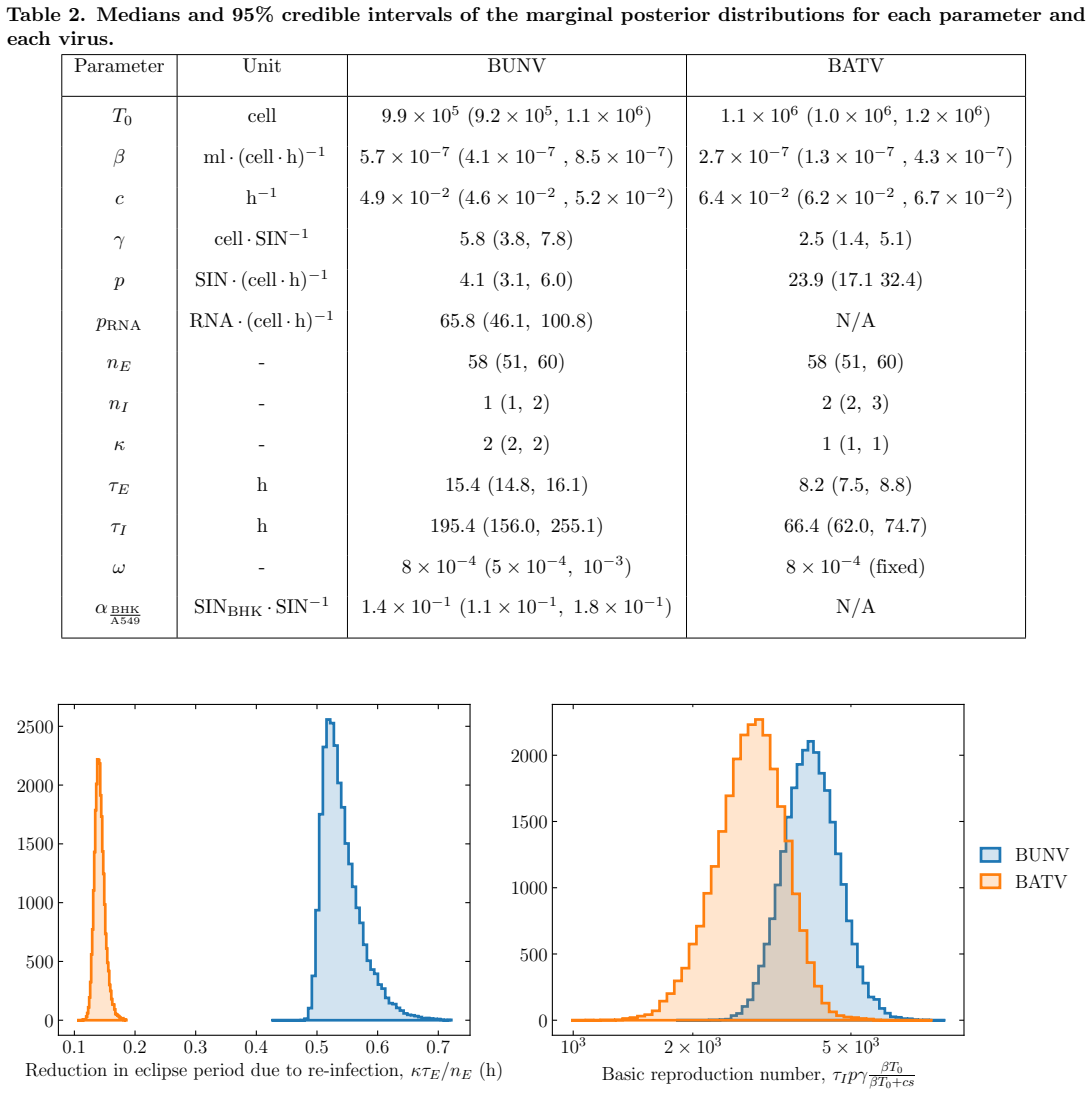
Bunyamwera virus has longer eclipse and infectious periods than Batai virus, with re-infection shortening the eclipse phase more strongly for BUNV.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
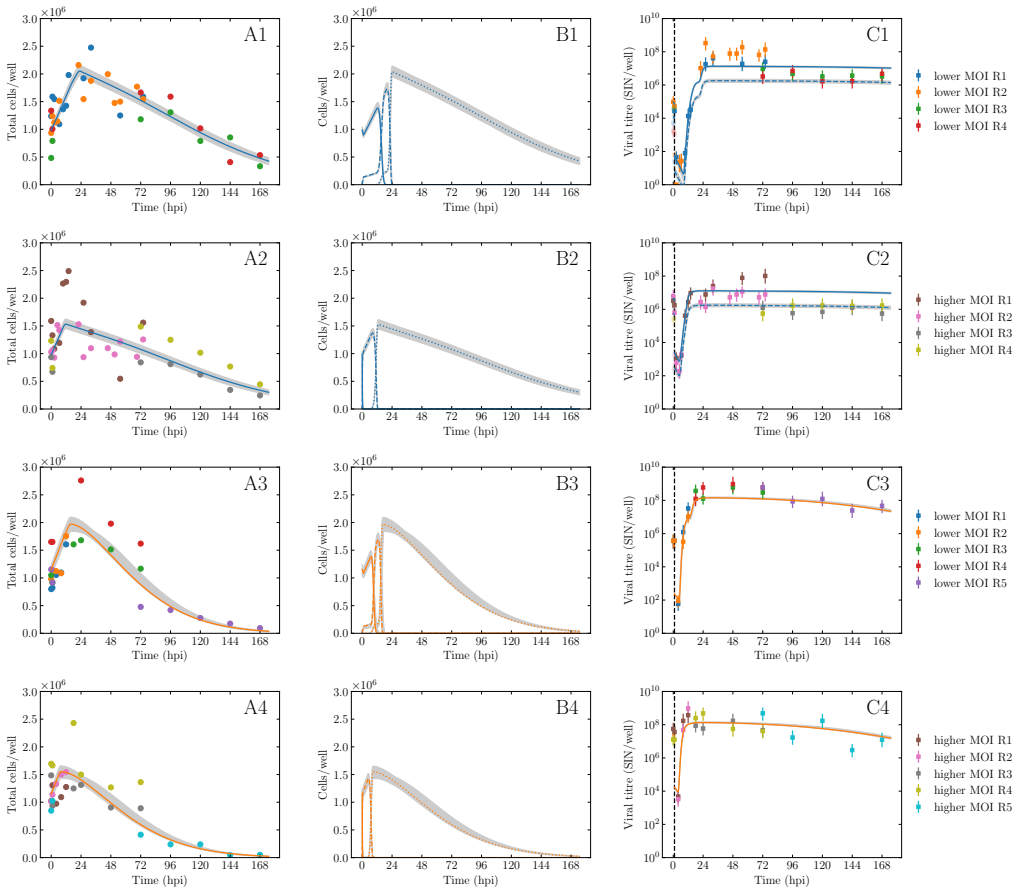
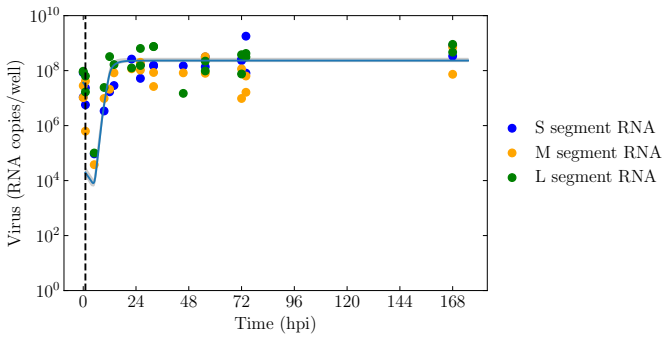
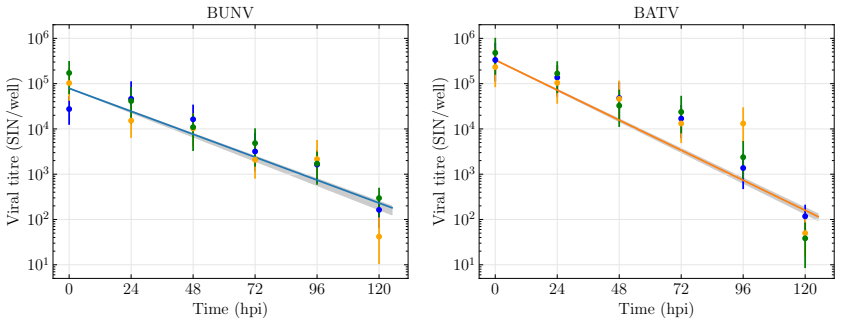
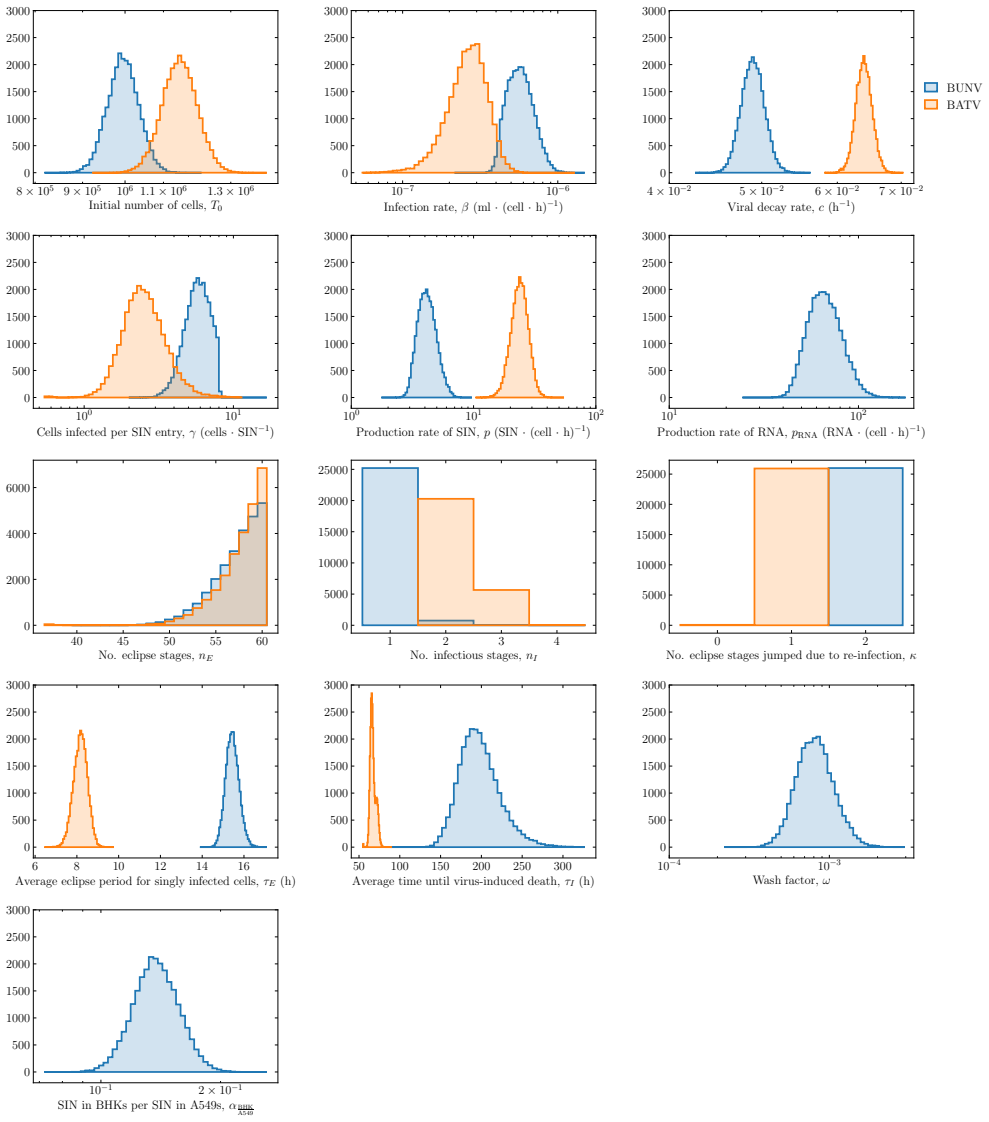
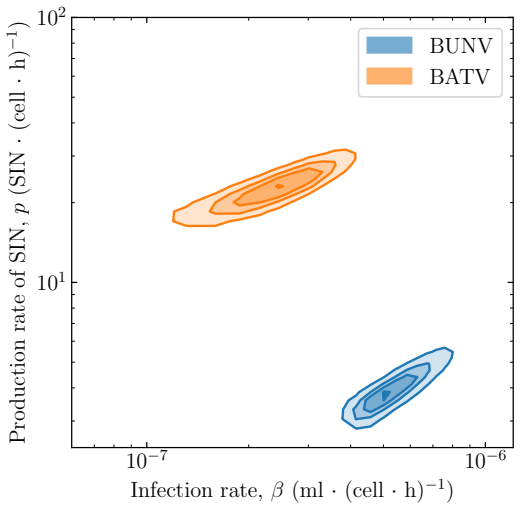
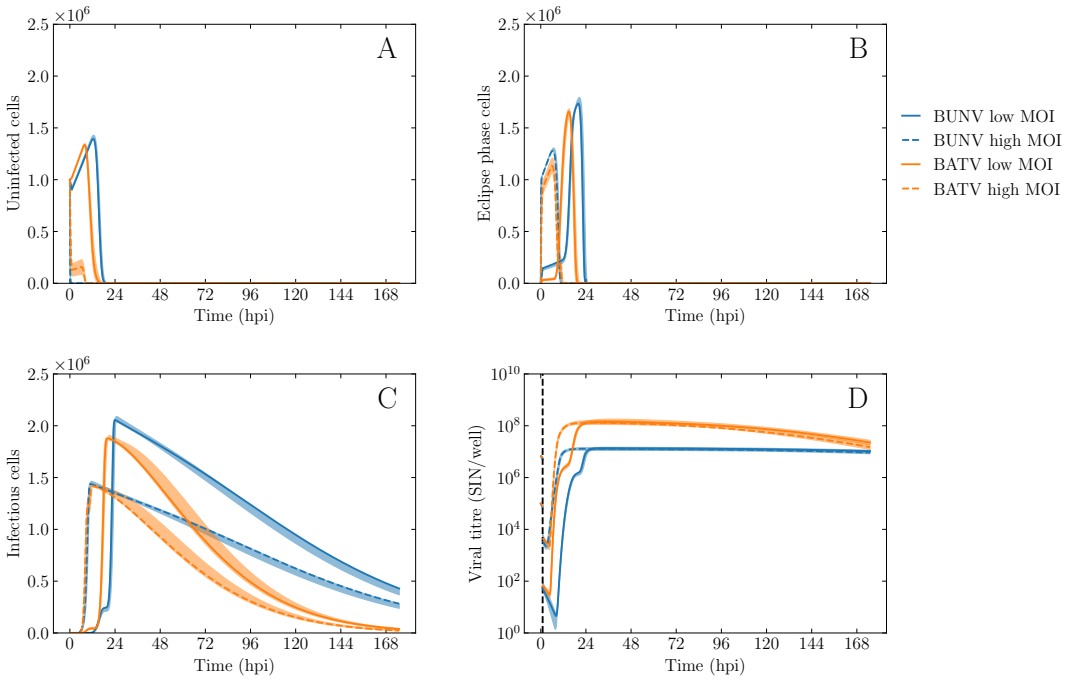
BUNV exhibited substantially longer eclipse and infectious periods than BATV, while BATV showed a higher per-cell virus production rate. Re-infection was predicted to shorten the eclipse phase for both viruses, but the effect was markedly stronger for BUNV. These results come from a model fit to decay data, growth curves, and genome copy measurements that quantifies the distinct in vitro viral kinetics of the two viruses.
What carries the argument
The deterministic model structure that incorporates MOI-dependent shortening of the eclipse phase due to re-infection, fitted via MCMC to constrain the per-infectious-unit infection rate.
If this is right
- BUNV has substantially longer eclipse and infectious periods than BATV.
- BATV produces virus at a higher rate per infected cell than BUNV.
- Re-infection shortens the eclipse phase more strongly for BUNV than for BATV.
- Genome copy measurements are essential to constrain the infection rate parameter for BUNV.
- The model distinguishes the replication dynamics of the two viruses quantitatively.
Where Pith is reading between the lines
- Differences in eclipse phase response to re-infection may affect how each virus spreads in insect vectors or mammalian hosts.
- Similar modeling could help compare kinetics of other bunyaviruses or related arboviruses.
- Direct measurement of eclipse phase at different MOIs could test the shortening prediction experimentally.
- The approach might inform models of within-host viral dynamics for these or similar pathogens.
Load-bearing premise
The MOI-dependent shortening of the eclipse phase is a genuine biological effect that the model structure can represent accurately.
What would settle it
Measuring the eclipse phase duration in cells infected at low MOI versus high MOI and finding no shortening for BUNV would falsify the re-infection effect.
Figures
read the original abstract
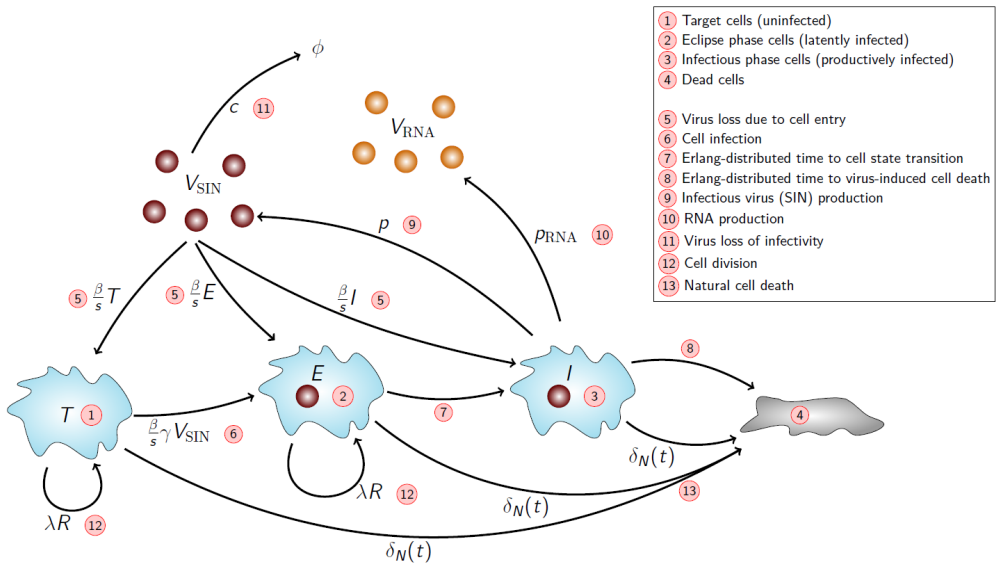
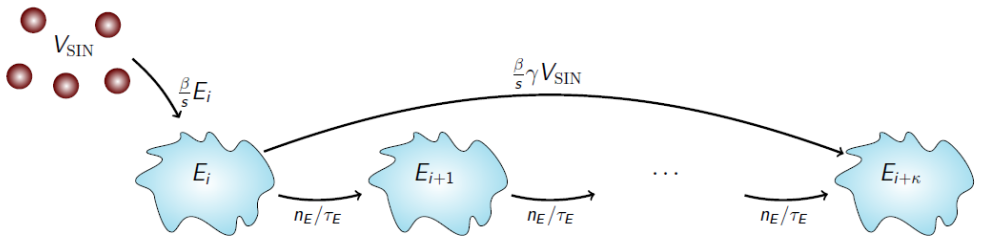
We develop a deterministic mathematical model to quantify the distinct in vitro infection dynamics of Bunyamwera virus (BUNV) and Batai virus (BATV) in A549 cells, incorporating cell division and natural death, continued entry of virions into already-infected cells, and shortening of the eclipse phase driven by re-infection. The model parameters were estimated making use of viral decay data, growth curves at two different inoculum concentrations, and extra-cellular genome copy measurements (for BUNV) via Markov chain Monte Carlo. Genome copy measurements were essential for constraining estimates of the number of cells that can become infected per unit of infectious virus for BUNV. We found that BUNV exhibited substantially longer eclipse and infectious periods than BATV, while BATV showed a higher per-cell virus production rate. Re-infection was predicted to shorten the eclipse phase for both viruses, but the effect was markedly stronger for BUNV. Together, these results provide a quantitative comparison of the in vitro viral kinetics of BUNV and BATV and reveal substantial differences in their replication dynamics.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
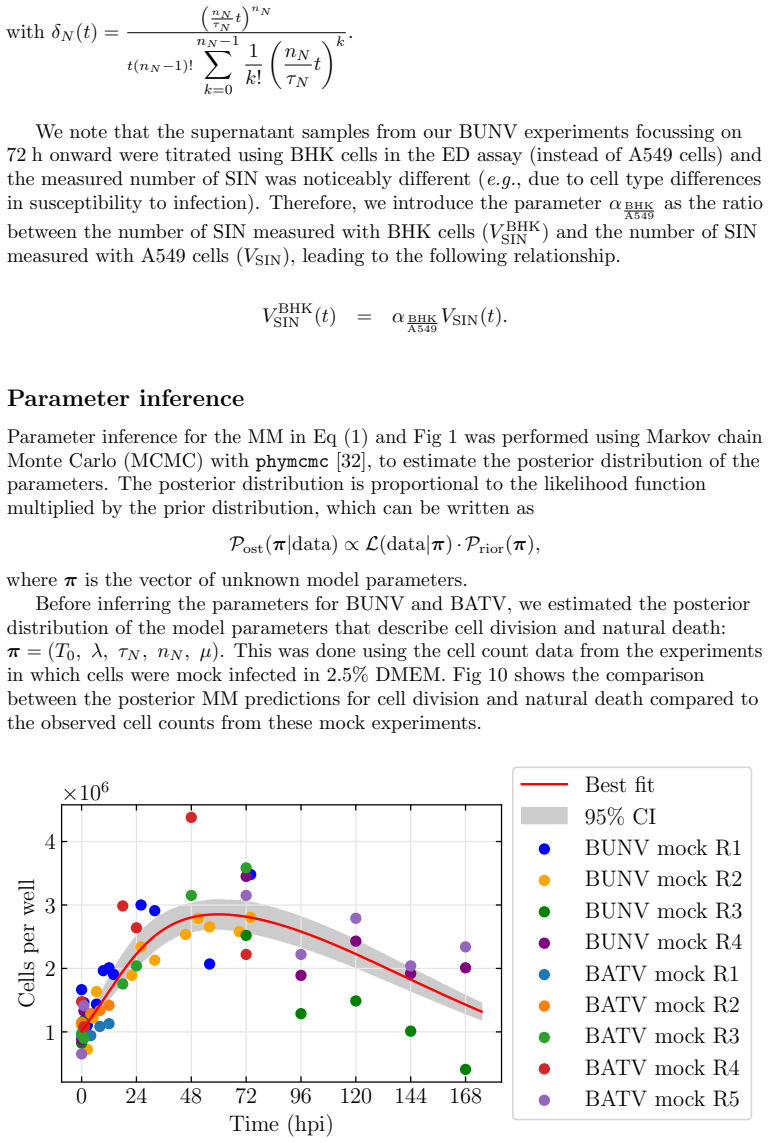
Summary. The paper develops a deterministic ODE model of in vitro BUNV and BATV infection in A549 cells that incorporates cell division and death, continued virion entry into infected cells, and linear shortening of the eclipse phase with cumulative re-infection events. Parameters are fit by MCMC to viral decay curves, growth curves at two MOIs, and (for BUNV only) extracellular genome-copy time series. The central claims are that BUNV has substantially longer eclipse and infectious periods than BATV, BATV has a higher per-cell production rate, and re-infection shortens the eclipse phase (more strongly for BUNV).
Significance. If the shortening term proves identifiable and necessary, the work supplies a quantitative, data-constrained comparison of two related bunyaviruses and demonstrates how re-infection can modulate eclipse duration in a virus-specific manner. The explicit use of genome-copy data to break a key degeneracy for BUNV is a methodological strength.
major comments (3)
- [Abstract] Abstract and Results: the reported shortening of the eclipse phase is an output of the MCMC fit to the same data used to estimate all other parameters rather than an independent prediction; consequently the claim that re-infection 'was predicted to shorten' the phase and that the effect is 'markedly stronger for BUNV' is circular and its magnitude is not falsifiable within the present analysis.
- [Methods] Methods/Results: no model-comparison statistics, Bayes factors, or profile-likelihood traces are shown to establish that the MOI-dependent shortening term is required by the data rather than being absorbed by compensatory changes in production rate or infectious-period length.
- [Results] Results: for BATV the degeneracy between the per-infectious-unit infection rate and eclipse duration is not broken by genome-copy data; without additional constraints or sensitivity analysis this undermines the reliability of the cross-virus comparison of eclipse-phase lengths.
minor comments (2)
- The full set of model equations, initial conditions, and the precise functional form of the eclipse-shortening term should be stated explicitly in the main text (or a dedicated supplementary section) rather than referenced only by name.
- Predictive-check figures should indicate which data types (decay, growth at MOI 1, growth at MOI 10, genome copies) are shown in each panel to allow readers to assess fit quality per data source.
Simulated Author's Rebuttal
We thank the referee for the constructive and detailed comments on our manuscript. We address each major comment below and indicate where revisions will be made to strengthen the presentation and analysis.
read point-by-point responses
-
Referee: [Abstract] Abstract and Results: the reported shortening of the eclipse phase is an output of the MCMC fit to the same data used to estimate all other parameters rather than an independent prediction; consequently the claim that re-infection 'was predicted to shorten' the phase and that the effect is 'markedly stronger for BUNV' is circular and its magnitude is not falsifiable within the present analysis.
Authors: We agree that the shortening effect is inferred from the MCMC posterior rather than constituting an independent prediction. The shortening term was included in the model on the basis of a mechanistic hypothesis regarding re-infection, and the data determine its estimated magnitude. To remove any implication of an out-of-sample prediction, we will revise the abstract and results sections to state that the model 'infers' or 'estimates' that re-infection shortens the eclipse phase (with a stronger effect for BUNV). This change makes the inferential nature of the result explicit. revision: yes
-
Referee: [Methods] Methods/Results: no model-comparison statistics, Bayes factors, or profile-likelihood traces are shown to establish that the MOI-dependent shortening term is required by the data rather than being absorbed by compensatory changes in production rate or infectious-period length.
Authors: The referee correctly notes the absence of formal model-comparison statistics. In the revised manuscript we will add a comparison of the full model against a reduced model lacking the MOI-dependent shortening term, reporting the deviance information criterion (DIC) from the MCMC runs and, where feasible, approximate Bayes factors. We will also inspect the joint posterior and profile likelihoods to confirm that the shortening parameter remains identifiable and is not fully compensated by adjustments in production rate or infectious period. revision: yes
-
Referee: [Results] Results: for BATV the degeneracy between the per-infectious-unit infection rate and eclipse duration is not broken by genome-copy data; without additional constraints or sensitivity analysis this undermines the reliability of the cross-virus comparison of eclipse-phase lengths.
Authors: We acknowledge that genome-copy data are available only for BUNV and that this leaves a potential degeneracy for BATV between infection rate and eclipse duration. The two-MOI growth curves supply some constraint via the timing of viral output, yet we agree that explicit sensitivity analysis is needed. We will add a sensitivity study for BATV in which the infection-rate parameter is varied over a plausible range while monitoring the resulting eclipse-duration posterior; the results will be reported to allow readers to assess the robustness of the cross-virus comparison. revision: partial
Circularity Check
Fitted re-infection shortening term presented as model prediction
specific steps
-
fitted input called prediction
[Abstract]
"Re-infection was predicted to shorten the eclipse phase for both viruses, but the effect was markedly stronger for BUNV."
The model structure incorporates 'shortening of the eclipse phase driven by re-infection' as an explicit term whose coefficient is estimated from the same viral growth and genome-copy datasets used to produce all other results. The reported shortening effect size is therefore the direct numerical output of that fitted coefficient rather than an independent prediction.
full rationale
The paper constructs a deterministic ODE model that explicitly includes an MOI-dependent shortening term for the eclipse phase, estimates all parameters (including the shortening coefficient) via MCMC on the growth curves and genome-copy data, and then reports the shortening as a 'prediction.' This is a fitted model feature rather than an out-of-sample or first-principles result, but the remaining parameter estimates (eclipse duration, infectious period, production rate) retain independent content from the data and are not forced by construction. No self-citation chains or definitional loops are present. The degeneracy noted by the skeptic is a statistical-identifiability issue, not circularity.
Axiom & Free-Parameter Ledger
free parameters (4)
- eclipse phase length
- infectious period length
- virus production rate per cell
- cells infected per infectious virion
axioms (2)
- standard math Infection dynamics follow deterministic ordinary differential equations
- domain assumption Cell division and natural death occur at constant rates in culture
Reference graph
Works this paper leans on
-
[1]
Mechanisms of bunyavirus morphogenesis and egress
Barker J, daSilva LL, Crump CM. Mechanisms of bunyavirus morphogenesis and egress. J Gen Virol. 2023;104(4):001845
2023
-
[2]
Recent advances in the molecular and cellular biology of bunyaviruses
Walter CT, Barr JN. Recent advances in the molecular and cellular biology of bunyaviruses. J Gen Virol. 2011;92(11):2467–2484
2011
-
[3]
Attenuation of bunyavirus replication by rearrangement of viral coding and noncoding sequences
Lowen AC, Boyd A, Fazakerley JK, Elliott RM. Attenuation of bunyavirus replication by rearrangement of viral coding and noncoding sequences. J Virol. 2005;79(11):6940–6946
2005
-
[4]
Visualizing the replication cycle of bunyamwera orthobunyavirus expressing fluorescent protein-tagged Gc glycoprotein
Shi X, van Mierlo JT, French A, Elliott RM. Visualizing the replication cycle of bunyamwera orthobunyavirus expressing fluorescent protein-tagged Gc glycoprotein. J Virol. 2010;84(17):8460–8469
2010
-
[5]
Key Golgi factors for structural and functional maturation of bunyamwera virus
Novoa RR, Calderita G, Cabezas P, Elliott RM, Risco C. Key Golgi factors for structural and functional maturation of bunyamwera virus. J Virol. 2005;79(17):10852–10863
2005
-
[6]
Requirement of the N-terminal region of orthobunyavirus nonstructural protein NSm for virus assembly and morphogenesis
Shi X, Kohl A, L´ eonard VH, Li P, McLees A, Elliott RM. Requirement of the N-terminal region of orthobunyavirus nonstructural protein NSm for virus assembly and morphogenesis. J Virol. 2006;80(16):8089–8099
2006
-
[7]
NSm is a critical determinant for bunyavirus transmission between vertebrate and mosquito hosts
Terhzaz S, Kerrigan D, Almire F, Szemiel AM, Hughes J, Parvy JP, et al. NSm is a critical determinant for bunyavirus transmission between vertebrate and mosquito hosts. Nat Commun. 2025;16(1):1214
2025
-
[8]
Innate Immune Response Against Batai Virus, Bunyamwera Virus, and Their Reassortants
Z¨ oller DD, S¨ aurich J, Metzger J, Jung K, Lepenies B, Becker SC. Innate Immune Response Against Batai Virus, Bunyamwera Virus, and Their Reassortants. Viruses. 2024;16(12):1833. June 23, 2026 24/26
2024
-
[9]
It’s in the mix: Reassortment of segmented viral genomes
Lowen AC. It’s in the mix: Reassortment of segmented viral genomes. PLOS Pathog. 2018;14(9):e1007200
2018
-
[10]
Batai and Ngari viruses: M segment reassortment and association with severe febrile disease outbreaks in East Africa
Briese T, Bird B, Kapoor V, Nichol ST, Lipkin WI. Batai and Ngari viruses: M segment reassortment and association with severe febrile disease outbreaks in East Africa. J Virol. 2006;80(11):5627–5630
2006
-
[11]
Comparative characterization of the reassortant Orthobunyavirus Ngari with putative parental viruses, Bunyamwera and Batai: in vitro characterization and ex vivo stability
Dutuze MF, Mayton EH, Macaluso JD, Christofferson RC. Comparative characterization of the reassortant Orthobunyavirus Ngari with putative parental viruses, Bunyamwera and Batai: in vitro characterization and ex vivo stability. J Gen Virol. 2021;102(2):001523
2021
-
[12]
Mammals preferred: Reassortment of Batai and Bunyamwera orthobunyavirus occurs in mammalian but not insect cells
Heitmann A, Gusmag F, Rathjens MG, Maurer M, Frankze K, Schicht S, et al. Mammals preferred: Reassortment of Batai and Bunyamwera orthobunyavirus occurs in mammalian but not insect cells. Viruses. 2021;13(9):1702
2021
-
[13]
Probing orthobunyavirus reassortment using Bunyamwera and Batai viruses as models
Bowen JM, Gunter K, Lunel AM, Omoga DC, Jones JE, Giesel H, et al. Probing orthobunyavirus reassortment using Bunyamwera and Batai viruses as models. PLOS Negl Trop Dis. 2025;19(5):e0013120
2025
-
[14]
Impact of the H275Y and I223V mutations in the neuraminidase of the 2009 pandemic influenza virus in vitro and evaluating experimental reproducibility
Paradis EG, Pinilla LT, Holder BP, Abed Y, Boivin G, Beauchemin CA. Impact of the H275Y and I223V mutations in the neuraminidase of the 2009 pandemic influenza virus in vitro and evaluating experimental reproducibility. PLOS One. 2015;10(5):e0126115
2009
-
[15]
(In)validating experimentally derived knowledge about influenza A defective interfering particles
Liao LE, Iwami S, Beauchemin CA. (In)validating experimentally derived knowledge about influenza A defective interfering particles. J R Soc Interface. 2016;13(124):20160412
2016
-
[16]
Uncovering critical properties of the human respiratory syncytial virus by combining in vitro assays and in silico analyses
Beauchemin CA, Kim YI, Yu Q, Ciaramella G, DeVincenzo JP. Uncovering critical properties of the human respiratory syncytial virus by combining in vitro assays and in silico analyses. PLOS One. 2019;14(4):e0214708
2019
-
[17]
Quantification of Ebola virus replication kinetics in vitro
Liao LE, Carruthers J, Smither SJ, Team CV, Weller SA, Williamson D, et al. Quantification of Ebola virus replication kinetics in vitro. PLOS Comput Biol. 2020;16(11):e1008375
2020
-
[18]
Cresta D, Warren DC, Quirouette C, Smith AP, Lane LC, Smith AM, et al. Time to revisit the endpoint dilution assay and to replace the TCID 50 as a measure of a virus sample’s infection concentration. PLOS Comput Biol. 2021;17(10):e1009480. doi:10.1371/journal.pcbi.1009480
-
[19]
Quirouette C, Thevakumaran R, Adachi K, Beauchemin CA. Does the random nature of cell-virus interactions during in vitro infections affect TCID 50 measurements and parameter estimation by mathematical models? arXiv preprint arXiv:241212960. 2024
2024
-
[20]
Incomplete bunyavirus particles can cooperatively support virus infection and spread
Berm´ udez-M´ endez E, Bronsvoort KF, Zwart MP, van de Water S, C´ ardenas-Rey I, Vloet RP, et al. Incomplete bunyavirus particles can cooperatively support virus infection and spread. PLOS Biol. 2022;20(11):e3001870
2022
-
[21]
Emerging infectious diseases: the Bunyaviridae
Soldan SS, Gonz´ alez-Scarano F. Emerging infectious diseases: the Bunyaviridae. Journal of neurovirology. 2005;11(5):412–423
2005
-
[22]
Tick-borne Bunyaviruses: an emerging public health threat; 2025
Hawman DW, Appelberg S, Duscher GG. Tick-borne Bunyaviruses: an emerging public health threat; 2025. June 23, 2026 25/26
2025
-
[23]
Oropouche virus: an emerging orthobunyavirus
Tilston-Lunel NL. Oropouche virus: an emerging orthobunyavirus. Journal of General Virology. 2024;105(10):002027
2024
-
[24]
Bunyamwera virus nonstructural protein NSs counteracts interferon regulatory factor 3-mediated induction of early cell death
Kohl A, Clayton RF, Weber F, Bridgen A, Randall RE, Elliott RM. Bunyamwera virus nonstructural protein NSs counteracts interferon regulatory factor 3-mediated induction of early cell death. J Virol. 2003;77(14):7999–8008
2003
-
[25]
Interaction of Bunyamwera Orthobunyavirus NSs protein with mediator protein MED8: a mechanism for inhibiting the interferon response
L´ eonard VH, Kohl A, Hart TJ, Elliott RM. Interaction of Bunyamwera Orthobunyavirus NSs protein with mediator protein MED8: a mechanism for inhibiting the interferon response. J Virol. 2006;80(19):9667–9675
2006
-
[26]
Lymphocytic choriomeningitis arenavirus utilises intercellular connections for cell to cell spread
Byford O, Shaw AB, Tse HN, Moon-Walker A, Saphire EO, Whelan SP, et al. Lymphocytic choriomeningitis arenavirus utilises intercellular connections for cell to cell spread. Sci Rep. 2024;14(1):28961
2024
-
[27]
Hepatitis C virus cell-cell transmission in hepatoma cells in the presence of neutralizing antibodies
Timpe JM, Stamataki Z, Jennings A, Hu K, Farquhar MJ, Harris HJ, et al. Hepatitis C virus cell-cell transmission in hepatoma cells in the presence of neutralizing antibodies. Hepatol. 2008;47(1):17–24
2008
-
[28]
Neutralizing antibody-resistant hepatitis C virus cell-to-cell transmission
Brimacombe CL, Grove J, Meredith LW, Hu K, Syder AJ, Flores MV, et al. Neutralizing antibody-resistant hepatitis C virus cell-to-cell transmission. J Virol. 2011;85(1):596–605
2011
-
[29]
Modulation of potassium channels inhibits bunyavirus infection
Hover S, King B, Hall B, Loundras EA, Taqi H, Daly J, et al. Modulation of potassium channels inhibits bunyavirus infection. J Biol Chem. 2016;291(7):3411–3422
2016
-
[30]
Bunyamwera orthobunyavirus glycoprotein precursor is processed by cellular signal peptidase and signal peptide peptidase
Shi X, Botting CH, Li P, Niglas M, Brennan B, Shirran SL, et al. Bunyamwera orthobunyavirus glycoprotein precursor is processed by cellular signal peptidase and signal peptide peptidase. Proc Natl Acad Sci. 2016;113(31):8825–8830
2016
-
[31]
Bunyaviruses: from transmission by arthropods to virus entry into the mammalian host first-target cells
L´ eger P, Lozach PY. Bunyaviruses: from transmission by arthropods to virus entry into the mammalian host first-target cells. Future Virol. 2015;10(7):859–881
2015
-
[32]
phymcmc: A convenient wrapper for emcee; 2019
Beauchemin CAA. phymcmc: A convenient wrapper for emcee; 2019. https://github.com/cbeauc/phymcmc. June 23, 2026 26/26
2019
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.