Soliton-like Waves in a Two-Dimensional Recurrent Spiking Neural Network with Weighted Spike-Timing-Dependent Plasticity
Pith reviewed 2026-06-26 12:46 UTC · model grok-4.3
The pith
A two-dimensional spiking neural network with weighted STDP spontaneously generates stable dissipative soliton waves from local rules.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
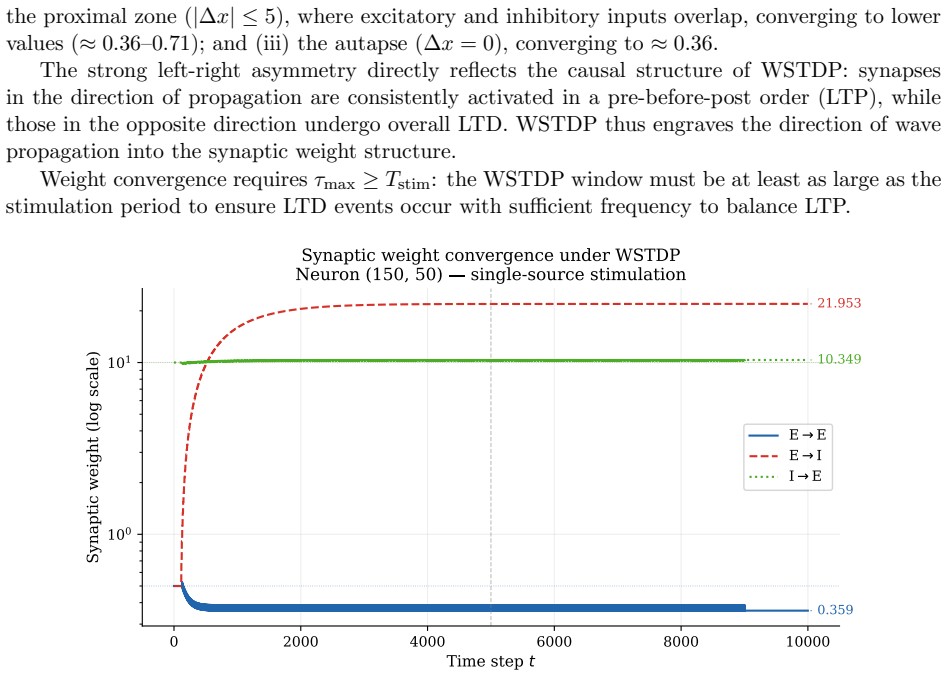
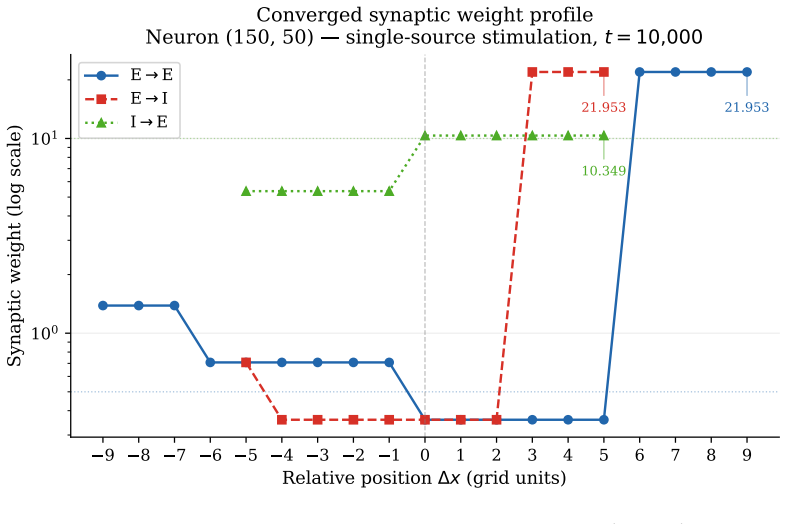
Assembling excitatory-inhibitory pairs of such neurons into a two-dimensional recurrent network and applying periodic localized stimulation, the network spontaneously gives rise to stable, self-propagating wave packets with the properties of dissipative solitons: they maintain a stable spatial profile, propagate at constant speed, and annihilate upon frontal collision. Their emergence requires a geometric asymmetry between excitatory and inhibitory connection radii, and initial inhibitory synapses stronger than excitatory ones. WSTDP engraves the direction of propagation into the synaptic weight profile, so that the network learns by itself to sustain propagation in one direction while suppr
What carries the argument
Two-dimensional recurrent network of excitatory-inhibitory neuron pairs with weighted spike-timing-dependent plasticity and asymmetric connection radii, which together produce spontaneous soliton-like waves and directional learning.
If this is right
- The waves maintain a stable spatial profile and travel at constant speed.
- Frontal collisions cause the waves to annihilate.
- WSTDP imprints unidirectional propagation into the synaptic weights.
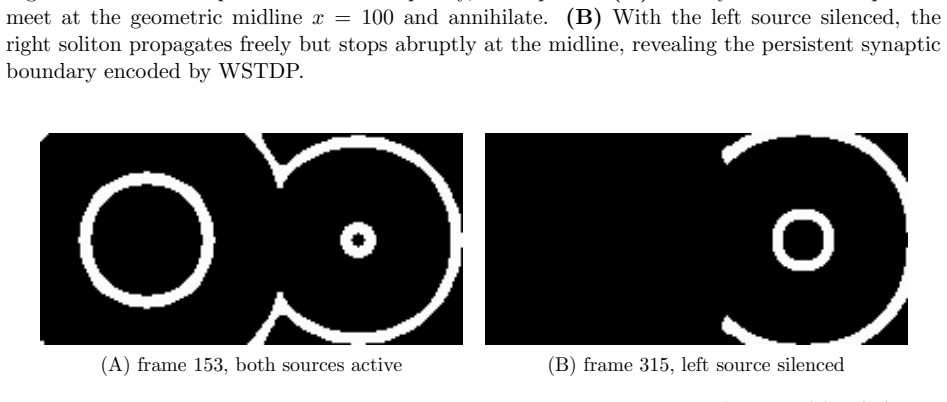
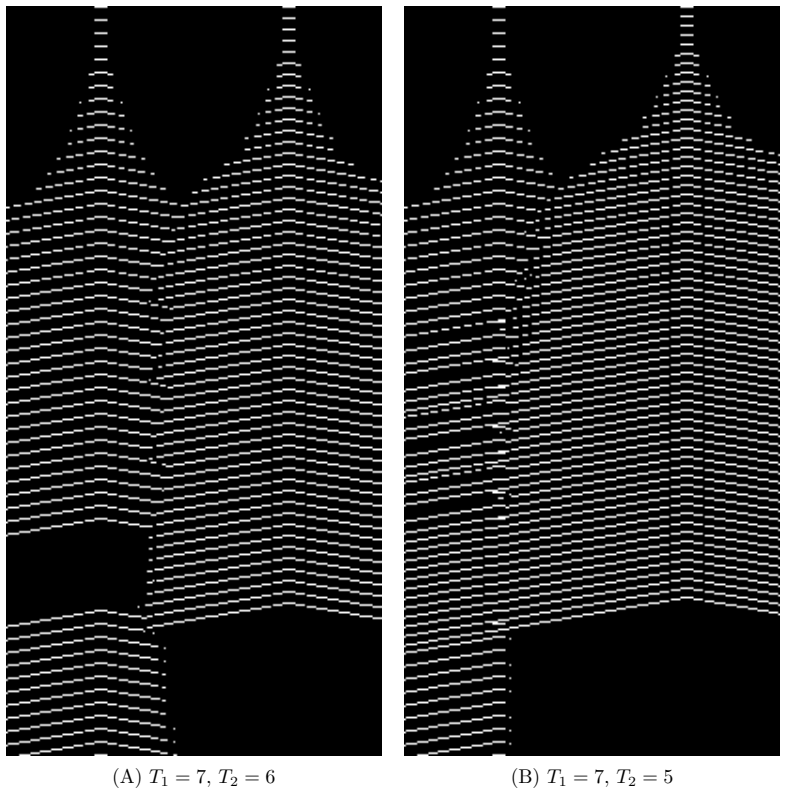
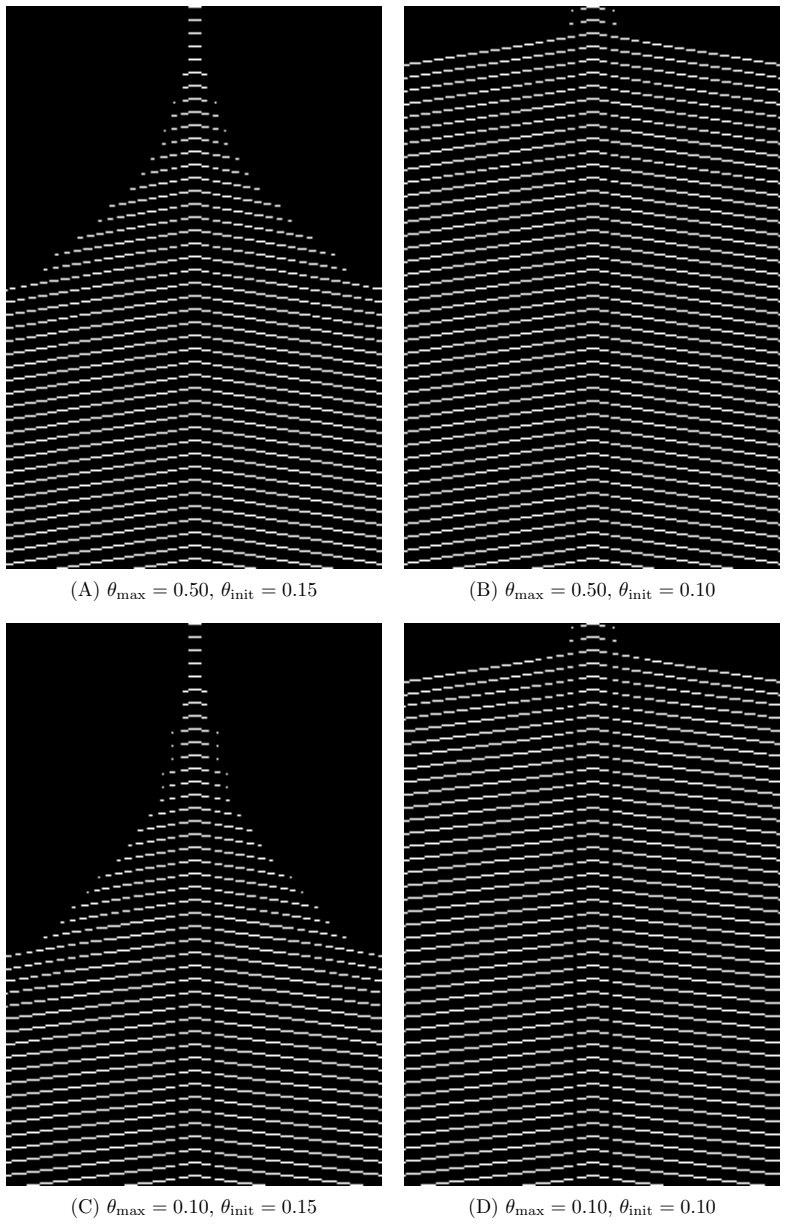
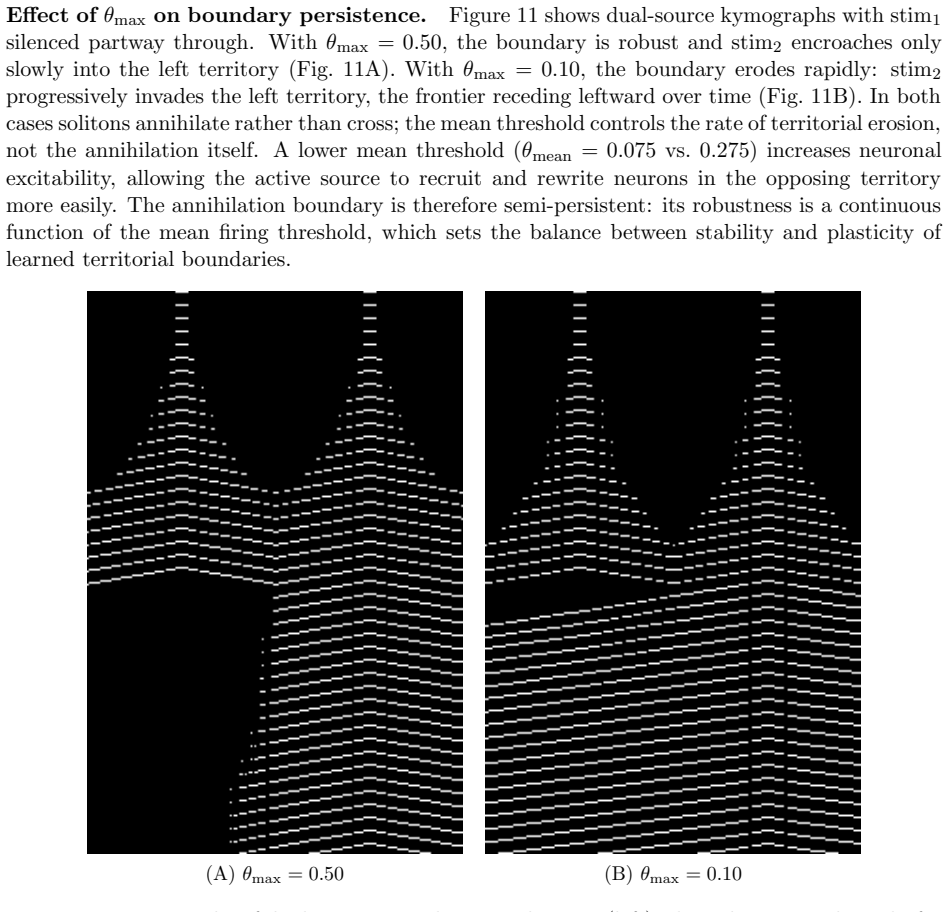
- Simultaneous activity from two sources produces a semi-persistent boundary whose location encodes relative phase and frequency.
- The construction supplies a minimal framework for cortical traveling waves, activity-zone boundaries, and spatial memory arising from local plasticity rules.
Where Pith is reading between the lines
- Similar local rules might allow biological cortex to generate and maintain directional traveling waves without external guidance or global signals.
- The collision-created boundary could function as a computational primitive for encoding spatial relations or timing differences in sensory processing.
- Adding realistic noise or scaling the network to larger sizes would test whether the soliton behavior persists beyond the minimal setting.
- The same asymmetry-plus-plasticity combination might be transplanted into other recurrent architectures to produce stable propagating patterns for computational tasks.
Load-bearing premise
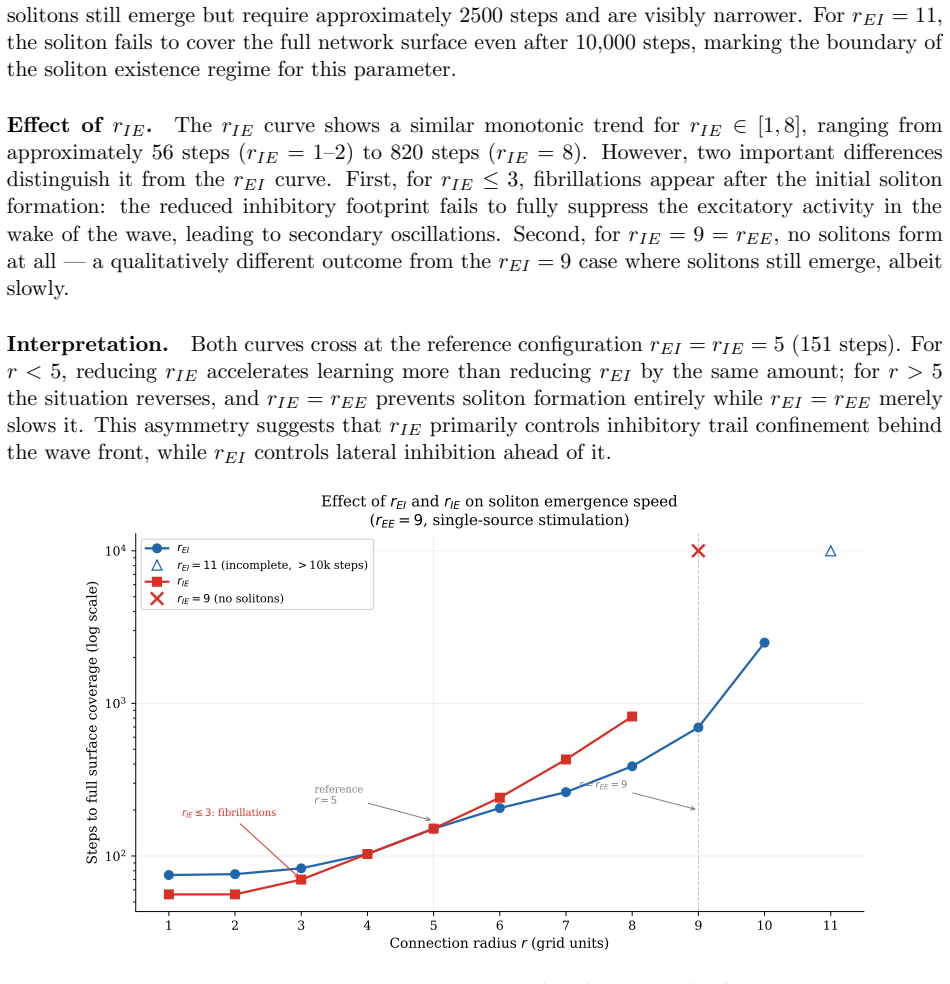
The emergence of these waves depends on a geometric asymmetry between the radii of excitatory and inhibitory connections together with initially stronger inhibitory synapses.
What would settle it
Simulating the network after removing the connection-radius asymmetry or setting initial excitatory and inhibitory synapse strengths equal, and finding no stable propagating waves, would falsify the necessity of those conditions.
Figures



read the original abstract
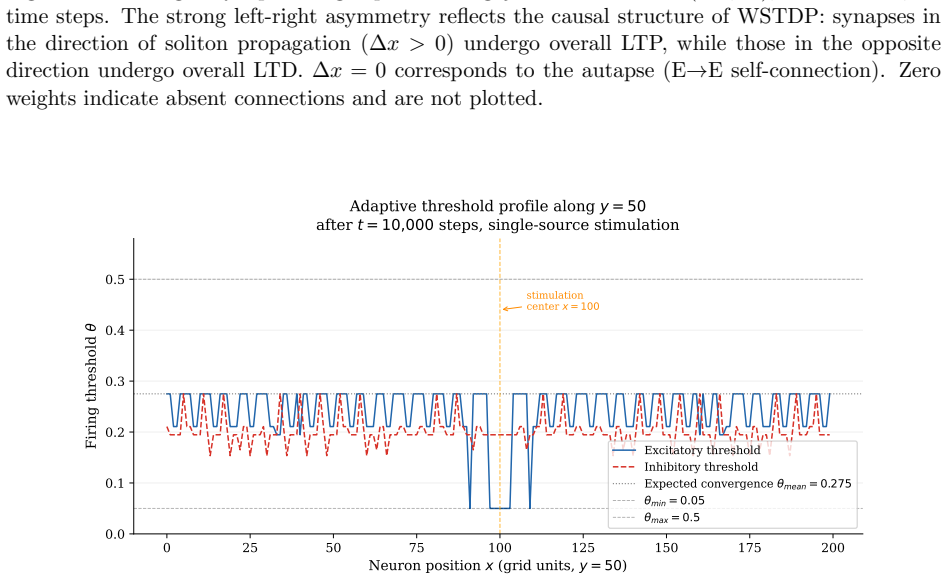
We construct a minimal but biologically plausible spiking neuron model operating in discrete time, combining multiplicative spike-timing-dependent plasticity (WSTDP), divisive normalization of synaptic integration, homeostatic threshold adaptation, and a one-step refractory period. We show that this normalization admits a biologically plausible dendritic implementation in which each binary junction operates using only locally available information. Assembling excitatory-inhibitory pairs of such neurons into a two-dimensional recurrent network and applying periodic localized stimulation, we find that the network spontaneously gives rise to stable, self-propagating wave packets with the properties of dissipative solitons: they maintain a stable spatial profile, propagate at constant speed, and annihilate upon frontal collision. Their emergence requires a geometric asymmetry between excitatory and inhibitory connection radii, and initial inhibitory synapses stronger than excitatory ones. WSTDP engraves the direction of propagation into the synaptic weight profile, so that the network learns by itself to sustain propagation in one direction while suppressing the reverse. When two sources are active simultaneously, the resulting waves annihilate upon collision, defining a semi-persistent boundary whose position encodes the relative phase and frequency of the two sources. These results provide a minimal computational framework for studying the emergence of cortical traveling waves, activity zone delimitation, and spatial memory from local plasticity rules alone.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript constructs a minimal discrete-time spiking neuron model combining weighted spike-timing-dependent plasticity (WSTDP), divisive normalization of synaptic integration (with a proposed local dendritic implementation), homeostatic threshold adaptation, and a one-step refractory period. Assembling excitatory-inhibitory neuron pairs into a 2D recurrent network with asymmetric excitatory versus inhibitory connection radii and stronger initial inhibitory synaptic weights, the authors apply periodic localized stimulation and report the spontaneous emergence of stable, self-propagating wave packets. These packets exhibit dissipative-soliton properties (stable spatial profile, constant propagation speed, frontal annihilation on collision), with WSTDP engraving propagation direction and multi-source collisions producing semi-persistent boundaries that encode relative phase and frequency.
Significance. If the reported numerical observations are robust, the work supplies a minimal, fully specified computational framework in which local, biologically motivated plasticity rules alone suffice to produce macroscopic traveling waves, direction learning, and activity-zone delimitation. The explicit disclosure of the two required asymmetries (connection radii and initial weight imbalance) and the dendritic normalization proposal are constructive features that facilitate reproducibility and biological interpretation.
minor comments (3)
- The manuscript should include a dedicated methods subsection (or appendix) tabulating all fixed parameter values, network size, stimulation protocol details, and the precise numerical criteria used to classify a wave packet as 'stable' and 'self-propagating' (e.g., profile correlation threshold, speed variance bound).
- Figure captions and main text should explicitly state the number of independent runs, random seeds, and any statistical tests confirming that the soliton properties persist across initial conditions once the required asymmetries are imposed.
- A brief comparison paragraph placing the observed wave speed and annihilation behavior against existing continuum or mean-field models of cortical traveling waves would help readers assess novelty.
Simulated Author's Rebuttal
We thank the referee for the detailed summary, positive significance assessment, and recommendation of minor revision. No major comments were listed in the report.
Circularity Check
No significant circularity detected
full rationale
The paper constructs a discrete-time spiking model from explicit local rules (multiplicative WSTDP, divisive normalization, homeostatic threshold, refractory period) and reports direct numerical outcomes under stated initial conditions (E-I radius asymmetry and stronger initial inhibitory weights). No equations are presented that derive a target quantity from a fitted parameter whose value is itself taken from the target; no self-citation chain is invoked to justify uniqueness or an ansatz; and the soliton-like properties are described as observed simulation results rather than predictions forced by construction. The derivation chain is therefore self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
free parameters (2)
- excitatory versus inhibitory connection radii
- initial inhibitory versus excitatory synaptic strengths
axioms (1)
- domain assumption The chosen normalization admits a biologically plausible dendritic implementation in which each binary junction operates using only locally available information.
Reference graph
Works this paper leans on
-
[1]
, title =
Amari, S. , title =. Biological Cybernetics , year =
-
[2]
Bennett, J. E. M. and Bair, W. , title =. PLOS Computational Biology , year =
-
[3]
Bi, G. Q. and Poo, M. M. , title =. Journal of Neuroscience , year =
-
[4]
Bressloff, P. C. , title =. Journal of Physics A: Mathematical and Theoretical , year =
-
[5]
Ermentrout, G. B. and Kleinfeld, D. , title =. Neuron , year =
-
[6]
Markram, H. and L\". Regulation of synaptic efficacy by coincidence of postsynaptic. Science , year =
-
[7]
and Thorpe, S
Masquelier, T. and Thorpe, S. J. , title =. PLOS Computational Biology , year =
-
[8]
and Diesmann, M
Morrison, A. and Diesmann, M. and Gerstner, W. , title =. Biological Cybernetics , year =
-
[9]
and Chavane, F
Muller, L. and Chavane, F. and Reynolds, J. and Sejnowski, T. J. , title =. Nature Reviews Neuroscience , year =
-
[10]
van Rossum, M. C. W. and Bi, G. Q. and Turrigiano, G. G. , title =. Journal of Neuroscience , year =
-
[11]
and Miller, K
Song, S. and Miller, K. D. and Abbott, L. F. , title =. Nature Neuroscience , year =
-
[12]
and Cruz, L
Butler, K. and Cruz, L. , title =. Journal of Computational Neuroscience , year =
-
[13]
Davis, Z. W. and Muller, L. and Martinez-Trujillo, J. and Sejnowski, T. and Reynolds, J. H. , title =. Nature , year =
-
[14]
and Inglebert, Y
Debanne, D. and Inglebert, Y. , title =. Current Opinion in Neurobiology , year =
-
[15]
and Garrido, J
Debanne, D. and Garrido, J. J. , title =. Encyclopedia of Life Sciences , publisher =. 2018 , doi =
2018
-
[16]
and Heeger, D
Carandini, M. and Heeger, D. J. , title =. Nature Reviews Neuroscience , year =
-
[17]
Kole, M. H. P. and Stuart, G. J. , title =. Neuron , year =
-
[18]
Trachtenberg, J. T. and Chen, B. E. and Knott, G. W. and Feng, G. and Sanes, J. R. and Welker, E. and Svoboda, K. , title =. Nature , year =
-
[19]
Douglas, R. J. and Martin, K. A. C. , title =. Annual Review of Neuroscience , year =
-
[20]
Laughlin, S. B. and Sejnowski, T. J. , title =. Science , year =
-
[21]
and Klausberger, T
Somogyi, P. and Klausberger, T. , title =. Journal of Physiology , year =
-
[22]
and Trappenberg, T
Standage, D. and Trappenberg, T. , title =. Proceedings of the 2007 International Joint Conference on Neural Networks (IJCNN) , pages =. 2007 , doi =
2007
-
[23]
Abraham, W. C. , title =. Nature Reviews Neuroscience , year =
-
[24]
Multimicroelectrode investigation of monkey striate cortex: spike train correlations in the infragranular layers , journal =
Kr\". Multimicroelectrode investigation of monkey striate cortex: spike train correlations in the infragranular layers , journal =. 1988 , volume =
1988
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.