Effect of swimming mode on shielding of odor traces in turbulence
Pith reviewed 2026-05-23 06:00 UTC · model grok-4.3
The pith
Pullerlike swimmers shield their odor traces more effectively than pusherlike swimmers in turbulent water.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Swimmers alter their odor plume shape through enhanced mixing from velocity fluctuations and swimmer-induced flow circulation that widens the plume close by while speeding dilution of the chemical trace. Pullerlike swimmers achieve greater olfactory shielding than pusherlike swimmers; the difference arises because dynamics at the swimmer location trap odor at the source for pushers and dilute it for pullers. The shielding effect is robust and more pronounced for weak turbulent Reynolds numbers and large swimmer Reynolds numbers.
What carries the argument
The local flow dynamics at the swimmer position, which trap odor for pusher models and dilute it for puller models during passive scalar advection in turbulence.
If this is right
- Swimming reduces the likelihood of odor detection at distances roughly ten times the group size or greater.
- The shielding benefit grows stronger when turbulent Reynolds number is low and swimmer Reynolds number is high.
- The difference between swimming modes may influence which styles persist in marine organisms.
- Short-range detection risk rises while long-range chemical traces weaken overall.
Where Pith is reading between the lines
- The source-region mechanism could influence how predators track prey or how groups exchange chemical signals in the ocean.
- Similar flow differences might affect transport of other scalars such as nutrients or heat around moving bodies.
- Laboratory experiments with dyed fluid around real microorganisms could directly test the predicted concentration patterns.
Load-bearing premise
Odor behaves as a passive scalar carried by the flow with no chemical reactions or sensing feedback, and the pusher and puller models correctly represent the far-field flow produced by real swimmers at the Reynolds numbers simulated.
What would settle it
Measurements of odor concentration fields immediately around living puller and pusher swimmers in a controlled turbulent flow would reveal whether the source-region dilution versus trapping difference actually occurs.
Figures
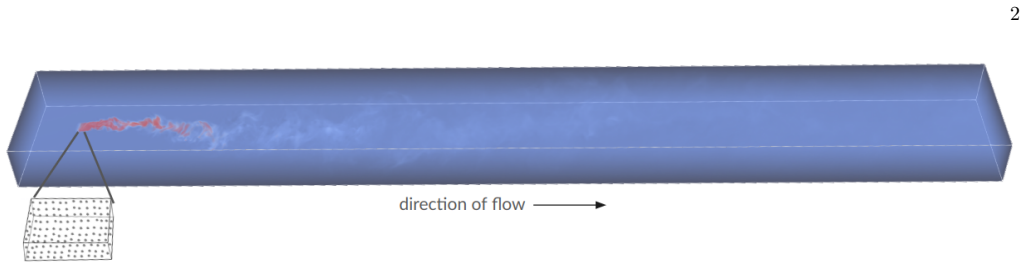




read the original abstract
Marine organisms manipulate their surrounding flow through their swimming dynamics, which affects the transport of their own odor cues. We demonstrate by direct numerical simulations how a group of swimmers, moving at intermediate Reynolds numbers, immersed in a turbulent flow, alter the shape of the odor plume they release in the water. Odor mixing is enhanced by increased velocity fluctuations and a swimmer-induced flow circulation that widens the odor plume at close range while speeding up dilution of the chemical trace. Beyond a short-range increase in the likelihood of being detected, swimming considerably reduces detections with effects that can persist at distances on the order of ten times the size of the group or more. We find that pullerlike swimmers are more effective at olfactory shielding than pusherlike swimmers. We trace this difference back to the dynamics at the swimmer location, which tends to trap odor at the source for pushers and to dilute it for pullers. Olfactory shielding is robust to changes in the conditions, and is more pronounced for weak turbulent Reynolds numbers and large swimmer Reynolds numbers. Our results suggest that olfactory shielding may play a role in the emergence of different swimming modalities by marine organisms.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript uses direct numerical simulations of groups of pusher- and puller-like swimmers at intermediate Reynolds numbers in turbulent flows, with odor modeled as a passive scalar. It claims that swimmer-induced flows enhance mixing, widen plumes at short range, and reduce long-range detections, with pullers providing superior olfactory shielding compared to pushers because local dynamics trap odor at the source for pushers but dilute it for pullers. Shielding is robust, stronger at low turbulent Re and high swimmer Re, and may explain evolutionary differences in swimming modes.
Significance. If the central distinction holds, the work identifies a hydrodynamic mechanism linking swimming modality to olfactory ecology in marine organisms. A strength is the use of forward DNS with no fitted parameters in the shielding metric, allowing controlled variation of swimmer type and turbulence strength to produce falsifiable plume statistics.
major comments (2)
- [Methods (swimmer forcing)] Methods (swimmer forcing implementation): the pusher/puller distinction relies on low-Re Stokes dipole solutions, yet simulations are performed at intermediate Re where inertial terms are non-negligible. No section quantifies the deviation of the DNS velocity field from the analytic far-field dipole at distances relevant to odor advection (e.g., several body lengths). This directly affects the load-bearing mechanistic claim in the results that the shielding difference traces to trapping versus dilution at the swimmer location.
- [Results (plume statistics)] Results (plume statistics and convergence): the central claim that pullers shield more effectively rests on reported differences in odor concentration fields and detection probabilities. No explicit demonstration of statistical convergence with respect to ensemble size, grid resolution, or domain size is referenced, which is required to establish that the reported superiority of pullers is not an artifact of under-resolved turbulence or swimmer wakes.
minor comments (1)
- [Abstract] Abstract: should include at least a brief statement on grid resolution, boundary conditions, and statistical convergence to allow readers to assess the reliability of the plume statistics without immediately consulting the methods.
Simulated Author's Rebuttal
We thank the referee for their careful reading of the manuscript and for the constructive comments. We address each major comment below and outline the revisions we will make.
read point-by-point responses
-
Referee: Methods (swimmer forcing implementation): the pusher/puller distinction relies on low-Re Stokes dipole solutions, yet simulations are performed at intermediate Re where inertial terms are non-negligible. No section quantifies the deviation of the DNS velocity field from the analytic far-field dipole at distances relevant to odor advection (e.g., several body lengths). This directly affects the load-bearing mechanistic claim in the results that the shielding difference traces to trapping versus dilution at the swimmer location.
Authors: We agree that the simulations operate at intermediate Reynolds numbers where inertial effects cause deviations from the low-Re Stokes dipole. The pusher/puller forcing follows the standard dipole implementation used in prior DNS studies of finite-Re swimmers. To directly support the mechanistic claim, we will add a new subsection quantifying the pointwise and far-field deviation between the DNS velocity and the analytic dipole at distances of several body lengths. This addition will be included in the revised Methods section. revision: yes
-
Referee: Results (plume statistics and convergence): the central claim that pullers shield more effectively rests on reported differences in odor concentration fields and detection probabilities. No explicit demonstration of statistical convergence with respect to ensemble size, grid resolution, or domain size is referenced, which is required to establish that the reported superiority of pullers is not an artifact of under-resolved turbulence or swimmer wakes.
Authors: We acknowledge that explicit convergence tests were not presented. The original simulations used resolutions and ensemble sizes consistent with established practices for resolving swimmer wakes and turbulent scalar transport, but we agree that a dedicated demonstration is needed. In the revision we will add an appendix with convergence checks for grid resolution, domain size, and ensemble size, supplemented by additional runs where necessary to confirm that the puller–pusher differences remain robust. revision: yes
Circularity Check
No circularity: results from forward DNS with imposed forcings
full rationale
The paper reports outcomes of direct numerical simulations solving the Navier-Stokes equations with passive scalar transport for odor. Pusherlike and pullerlike modes are imposed by the choice of swimmer forcing term in the setup; shielding metrics (plume shape, detection likelihood, dilution) are computed directly from the simulated fields. No equations, parameters, or metrics are defined in terms of the target results, and no load-bearing step reduces by construction to a fit, self-citation chain, or ansatz smuggled from prior work. The derivation chain is therefore self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
free parameters (2)
- swimmer Reynolds number
- turbulent Reynolds number
axioms (2)
- domain assumption Odor concentration obeys the advection-diffusion equation with no reaction terms.
- domain assumption Swimmers are modeled as fixed force dipoles (pusher or puller) without self-propulsion feedback on the flow.
Reference graph
Works this paper leans on
-
[1]
D.R. Webster and M.J. Weissburg. The hydrodynamics of chemical cues among aquatic organisms. Annu. Rev. Fluid Mech., 41:73–90, 2009
work page 2009
-
[2]
G. Reddy, V.N. Murthy, and M. Vergassola. Olfactory sensing and navigation in turbulent environments. Annu. Rev. Condens. Matter Phys. , 13:191–213, 2022
work page 2022
-
[3]
Biologically generated mixing in the ocean
Eric Kunze. Biologically generated mixing in the ocean. Annual review of marine science , 11(1):215–226, 2019
work page 2019
-
[4]
A viscosity- enhanced mechanism for biogenic ocean mixing
Kakani Katija and John O Dabiri. A viscosity- enhanced mechanism for biogenic ocean mixing. Nature, 460(7255):624–626, 2009
work page 2009
-
[5]
Biomixing of the oceans? Science, 316(5826):838–839, 2007
Andr´ e W Visser. Biomixing of the oceans? Science, 316(5826):838–839, 2007
work page 2007
-
[6]
S.K. Delavan and D.R. Webster. Predator and flow in- fluence on bivalve clam excurrent jet characteristics. J. Exp. Mar. Biol. Ecol. , 432:1–8, 2012
work page 2012
-
[7]
A. Alvarez. Modeling the odor-landscape resulting from the pumping behavior of bivalve clams in the presence of predators. J. Theor. Biol. , 453:40–47, 2018
work page 2018
- [8]
-
[9]
Fish swimming and bird/insect flight
Theodore Yaotsu Wu. Fish swimming and bird/insect flight. Annual review of fluid mechanics , 43(1):25–58, 2011
work page 2011
-
[10]
L. Ratnarajah, R. Abu-Alhaija, A. Atkinson, S. Batten, N.J. Bax, K.S. Bernard, G. Canonico, A. Cornils, J.D. Everett, M. Grigoratou, et al. Monitoring and modelling marine zooplankton in a changing climate. Nat. Com- mun., 14(1):564, 2023
work page 2023
-
[11]
G.R. Flierl and N.W. Woods. Copepod aggregations: influences of physics and collective behavior. J. Stat. Phys., 158:665–698, 2015
work page 2015
-
[12]
F.-G. Michalec, S. Souissi, and M. Holzner. Turbu- lence triggers vigorous swimming but hinders motion strategy in planktonic copepods. J. R. Soc. Interface , 12(106):20150158, 2015
work page 2015
-
[13]
M. Omori and W.M. Hamner. Patchy distribution of zoo- plankton: behavior, population assessment and sampling problems. Mar. Biol., 72:193–200, 1982
work page 1982
- [14]
-
[15]
The hydrodynam- ics of swimming microorganisms
Eric Lauga and Thomas R Powers. The hydrodynam- ics of swimming microorganisms. Reports on progress in physics, 72(9):096601, 2009
work page 2009
-
[16]
T. Kiørboe, H. Jiang, R.J. Gon¸ calves, L.T. Nielsen, and N. Wadhwa. Flow disturbances generated by feeding and swimming zooplankton. Proc. Natl. Acad. Sci. U.S.A. , 111(32):11738–11743, 2014
work page 2014
-
[17]
L.A. van Duren and J.J. Videler. Escape from viscos- ity: the kinematics and hydrodynamics of copepod forag- ing and escape swimming. J. Exp. Biol., 206(2):269–279, 2003
work page 2003
-
[18]
Arne W Zantop and Holger Stark. Emergent collective dynamics of pusher and puller squirmer rods: swarming, clustering, and turbulence. Soft Matter , 18(33):6179– 6191, 2022
work page 2022
-
[19]
Swarm of slender pusher and puller swim- mers at finite reynolds numbers
M Cavaiola. Swarm of slender pusher and puller swim- mers at finite reynolds numbers. Physics of Fluids, 34(2), 2022
work page 2022
- [20]
-
[21]
M. Quadrio, B. Frohnapfel, and Y. Hasegawa. Does the choice of the forcing term affect flow statistics in dns of turbulent channel flow? Eur. J. Mech. B Fluids , 55:286– 293, 2016
work page 2016
-
[22]
N. Rigolli, N. Magnoli, L. Rosasco, and A. Seminara. Learning to predict target location with turbulent odor plumes. Elife, 11:e72196, 2022
work page 2022
-
[23]
E. Selander, S.T. Fredriksson, and L. Arneborg. Chem- ical signaling in the turbulent ocean—hide and seek at the kolmogorov scale. Fluids, 5(2):54, 2020
work page 2020
-
[24]
H. Jiang and T.R. Osborn. Hydrodynamics of copepods: a review. Surv. Geophys., 25:339–370, 2004
work page 2004
-
[25]
J. Drescher, K.and Dunkel, L.H. Cisneros, S. Ganguly, and R.E. Goldstein. Fluid dynamics and noise in bacte- rial cell–cell and cell–surface scattering. Proc. Natl. Acad. Sci. U.S.A., 108(27):10940–10945, 2011
work page 2011
-
[26]
G.L. Wagner, W.R. Young, and E. Lauga. Mixing by mi- croorganisms in stratified fluids. J. Mar. Res., 72(2):47– 8 72, 2014
work page 2014
-
[27]
B. Vent´ ejou, T. M´ etivet, A. Dupont, and P. Peyla. Uni- versal scaling laws for a generic swimmer model. arXiv preprint arXiv:2407.04511, 2024
-
[28]
T.C. Granata and T.D. Dickey. The fluid mechanics of copepod feeding in a turbulent flow: a theoretical ap- proach. Prog. Oceanogr., 26(3):243–261, 1991
work page 1991
-
[29]
S Wang and A Ardekani. Inertial squirmer. Physics of Fluids, 24(10), 2012. 1 Supporting Information: Swimming mode determines how well mesoscale swimmers shield their odor in turbulence I. MODELING SWIMMER DYNAMICS As noted in the main text, swimmers are modeled as force dipoles oriented along the streamwise direction, repre- sented as F (δ(x + r) − δ(x −...
work page 2012
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.